Forskningsområde
Den stora majoriteten av de celler som är involverade i däggdjursimmunitet härstammar från prekursorer i benmärgen (vänstra halvan av figur 1) och cirkulerar i blodet och kommer in i och ibland ut ur vävnaderna när det behövs. En mycket sällsynt stamcell finns kvar i den vuxna benmärgen (med en frekvens på cirka 1 på 100 000 celler) och behåller förmågan att differentiera sig till alla typer av blodceller. Blodbildningen har studerats antingen genom att injicera små mängder genetiskt märkta märgceller i mottagande möss och observera vilken avkomma de ger upphov till (kloning in vivo) eller genom att odla benmärgsprekursorer i närvaro av lämpliga tillväxtfaktorer (kloning in vitro). Proliferation och differentiering av alla dessa celler kontrolleras av lösliga eller membranbundna tillväxtfaktorer som produceras av benmärgsstroma och av varandra. I cellen aktiverar dessa signaler specifika transkriptionsfaktorer, DNA-bindande molekyler som fungerar som huvudbrytare som bestämmer det efterföljande genetiska programmet, vilket i sin tur ger upphov till utvecklingen av de olika celltyperna (så kallade linjesträckningar). Anmärkningsvärt nog har nyligen genomförda studier visat att det är möjligt att omvandla en differentierad celltyp till en annan genom att experimentellt införa rätt transkriptionsfaktorer i cellen. Denna upptäckt har viktiga terapeutiska konsekvenser, t.ex. när det gäller att bota genetiska immunbrister. De flesta blodbildande celler slutar att dela sig när de är helt differentierade. Lymfocyter delar sig dock snabbt och expanderar efter exponering för antigen. Det ökade antalet lymfocyter som är specifika för ett antigen utgör grunden för det immunologiska minnet.
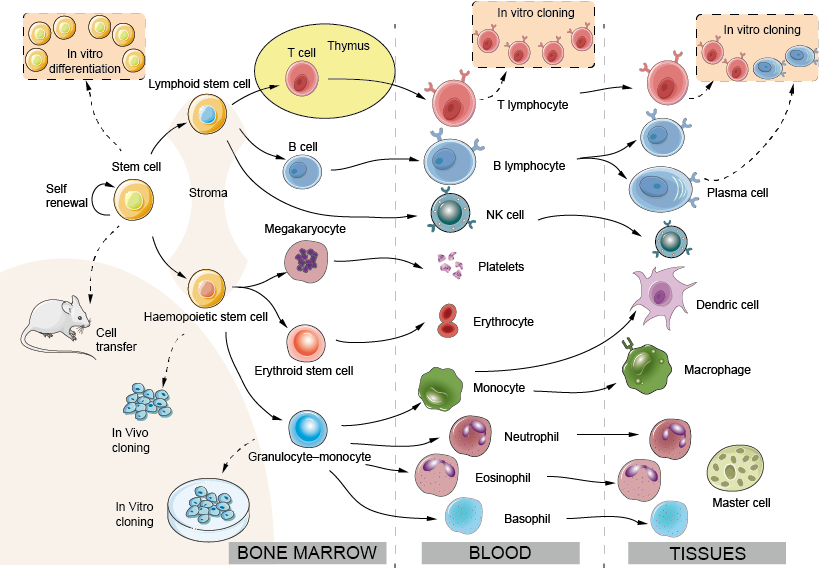
Figur 1. Immuncellsutveckling: det hemopoetiska systemet
Benskärl: Till skillnad från de flesta andra vävnader eller organ förnyar sig det hemopoetiska systemet ständigt. Hos vuxna sker utvecklingen av hemopoetiska celler huvudsakligen i benmärgen. Hos fostret, innan benen utvecklas, sker hemopoetisk cellbildning först i gulesäcken och sedan i levern.
Stroma: Epitel- och endotelceller som ger stöd och utsöndrar tillväxtfaktorer för hemopoetisk cellbildning.
Stamcell: Den totipotenta och självförnyande märgcellen. Stamceller finns i lågt antal i såväl blod som benmärg och antalet kan ökas genom behandling med lämpliga tillväxtfaktorer (t.ex. G-CSF), vilket underlättar benmärgstransplantationsprocessen avsevärt.
Lymfoid stamcell: antas kunna differentieras till T- eller B-lymfocyter. Mycket färska uppgifter tyder på att skillnaden mellan lymfoida och myeloida stamceller i själva verket kan vara mer komplex.
Hemopoetisk stamcell: föregångare till mjältknutor och kan troligen differentiera sig till alla utom de lymfoida banorna, dvs. granulocyter, erytroider, monocyter och megakaryocyter; kallas ofta CFU-GEMM.
Erytroid stamcell: ger upphov till erytrocyter. Erytropoietin, ett glykoproteinhormon som bildas i njuren som svar på hypoxi, påskyndar differentieringen av prekursorer av röda blodkroppar och anpassar på så sätt produktionen av röda blodkroppar till efterfrågan på deras syrebärande kapacitet, ett typiskt exempel på ”negativ återkoppling”.
Granulocyt-monocyt-gemensam prekursor: Den relativa andelen av dessa två celltyper regleras av ”tillväxt-” eller ”kolonistimulerande” faktorer.
Cloning: Man har undersökt enskilda stamcellers potential att ge upphov till en eller flera typer av hemopoetiska celler genom att isolera enskilda celler och låta dem dela sig många gånger och sedan observera vilka celltyper som återfinns bland avkomman. Denna process är känd som kloning (en klon är en uppsättning dotterceller som alla härstammar från en enda föräldracell). Bevis tyder på att en enda stamcell under vissa förhållanden kan ge upphov till alla fullt differentierade celler i ett vuxet hemopoetiskt system.
Neutrofil (polymorf): Den vanligaste leukocyten i människans blod, en kortlivad fagocytisk cell vars granula innehåller många bakteriedödande ämnen. Neutrofilerna är de första cellerna som lämnar blodet och går in på infektions- eller inflammationsställen.
Eosinofil: En leukocyt med stora refraktila granuler som innehåller ett antal mycket basiska eller ”katjoniska” proteiner, som möjligen är viktiga för att döda större parasiter, bland annat maskar.
Basofil: En leukocyt med stora basofila granuler som innehåller heparin och vasoaktiva aminer, som är viktiga i inflammationsreaktionen. De tre ovanstående celltyperna kallas ofta tillsammans för ”granulocyter”.
Megakaryocyt: Modercell till blodplättarna.
Blodplättar: Små celler som ansvarar för att försegla skadade blodkärl (”hemostas”) men som också är källan till många inflammatoriska mediatorer.
Monocyt: En prekursorcell i blodet som utvecklas till en makrofag när den migrerar in i vävnaderna. Ytterligare monocyter attraheras till inflammationsställen, vilket ger en reservoar av makrofager och kanske även dendritiska celler.
Makrofag: Den viktigaste inhemska fagocyten i vävnader och serösa håligheter såsom pleura och peritoneum.
DC (dendritisk cell): Dendritiska celler: Dendritiska celler finns i alla kroppens vävnader (t.ex. de Langerhanska cellerna i huden) där de tar upp antigen och sedan migrerar till T-cellområdena i lymfkörteln eller mjälten via lymfkärlen eller blodet. Deras viktigaste funktion är att aktivera T-cellsimmunitet, men de kan också vara inblandade i toleransinduktion. En andra undergrupp av plasmacytoida DC (ett namn som härrör från deras morfologiska likhet med plasmaceller) är de viktigaste producenterna av typ I-interferoner, en viktig grupp antivirala proteiner. Även om dendritiska celler experimentellt sett ofta härstammar från myeloida celler, är den dendritiska cellens utvecklingslinje i benmärgen fortfarande föremål för debatt.
NK-cell (natural killer): En lymfocytliknande cell som kan döda vissa virusinfekterade celler och vissa tumörceller, men med komplexa receptoruppsättningar som är helt skilda från de som finns på riktiga lymfocyter (för mer information se fig. 10). NK-celler och T-celler kan ha en gemensam föregångare.
T- och B-lymfocyter: T-lymfocyter (från thymus) och B-lymfocyter (från benmärg eller, hos fåglar, från bursa) är de viktigaste cellulära komponenterna i den adaptiva immuniteten. B-lymfocyter är föregångare till antikroppsbildande celler. Under fosterlivet kan levern spela rollen som ”bursa”.
Plasmacell: En B-cell i sitt höghastighetsantikroppsutsöndrande tillstånd. Trots sitt namn ses plasmaceller sällan i blodet, men finns i mjälte, lymfkörtlar etc. när antikroppar tillverkas. Plasmaceller delar sig inte och kan inte upprätthållas under längre perioder in vitro. B-lymfocyter som producerar specifika antikroppar kan dock fusioneras med en tumörcell för att producera en odödlig hybridklon eller ”hybridom”, som fortsätter att utsöndra antikroppar av en förutbestämd specificitet. Sådana monoklonala antikroppar har visat sig vara av enormt värde som specifika verktyg inom många grenar av biologin, och flera av dem används nu rutinmässigt för behandling av autoimmuna sjukdomar och cancer.
Mastcell: En stor vävnadscell som härrör från den cirkulerande basofilen. Mastceller utlöses snabbt av vävnadsskador för att initiera det inflammatoriska svar som orsakar många former av allergi.
Växtfaktorer: De molekyler som styr blodbildande cellers proliferation och differentiering är ofta också involverade i regleringen av immunsvar – interleukiner eller cytokiner. Vissa av dem upptäcktes först av hematologer och kallas ”kolonistimulerande faktorer” (CSF), men de olika namnen har ingen egentlig betydelse, och en av dem, IL-3, är faktiskt ofta känd som ”multi-CSF”. Tillväxtfaktorer används i klinisk praxis för att öka vissa undergrupper av blodkroppar, och erytropoietin var en av de första av den nya generationens proteiner som framställdes med hjälp av ”rekombinant” teknik och som användes i kliniken, och även av idrottsmän som ville öka antalet röda blodkroppar.
Leave a Reply