Frontiers in Physiology
Introducere
Anemia este una dintre cele mai frecvente afecțiuni la nivel mondial, iar anemia datorată deficienței de fier este forma prevalentă conform mai multor analize (recenzie în Camaschella, 2019). Acest tip de anemie rezultă din deficitul total de fier din organism și din incapacitatea de a furniza cantitatea mare de fier pe care măduva osoasă o consumă pentru a produce un număr adecvat de globule roșii pentru a menține oxigenarea țesuturilor.
Disponibilitatea fierului este controlată de hormonul peptidic hepatic hepcidină. Creșterea cantității de fier din organism determină producerea de hepcidină, care este eliberată în circulație și acționează asupra receptorului său, feroportina, o proteină transmembranară exportatoare de fier foarte exprimată pe enterocite, macrofage și hepatocite. Hepcidina reduce intrarea fierului în plasmă de la celulele duodenale absorbante și macrofagele care reciclează fierul prin blocarea exportului de fier (Aschemeyer et al., 2018) și prin degradarea exportatorului de fier feroportina (Nemeth et al., 2004). Prin reglarea fierului plasmatic și a homeostaziei sistemice a fierului, axa hepcidină/ferroportină afectează puternic eritropoieza, de unde și posibila apariție a anemiei.
The Iron-Erythropoiesis Connection
Procesul de producere a globulelor roșii consumă aproximativ 80% din fierul circulant pentru sinteza hemoglobinei eritroblastelor aflate la maturitate. Cea mai mare parte a fierului (20-25 mg/zi) este reciclat de către macrofage, în timp ce o cantitate limitată (1-2 mg zilnic) provine din absorbția intestinală. Hormonul renal eritropoietină (EPO) controlează proliferarea progenitorilor eritroizi, în special a CFU-e și, într-un grad mai redus, a BFU-e, și faza timpurie a eritropoiezei terminale, în timp ce nevoile de fier sunt crescute în etapele târzii de diferențiere de la proeritroblaste la reticulocit, pentru sinteza de heme încorporat în hemoglobină (Muckenthaler et al., 2017).
Reglarea hepcidinei necesită o legătură încrucișată între celulele sinusoidale endoteliale hepatice (LSEC) care produc proteinele morfogenetice osoase (BMP) pentru a activa calea BMP-SMAD și hepatocitele care produc și eliberează hepcidină (Babitt et al., 2006; Rausa et al., 2015). BMP6 și BMP2 sunt cele mai importante BMP care stimulează hepcidina, în timp ce expresia BMP6 este dependentă de fier (Andriopoulos et al., 2009; Meynard et al., 2009) BMP2 pare să fie mai puțin sensibilă la fier (Canali et al., 2017; Koch et al., 2017).
Nivelurile de hepcidină sunt scăzute în deficitul absolut de fier și în anemia feriprivă. În aceste condiții, depozitele de fier sunt epuizate și semnalizarea BMP-SMAD este dezactivată la mai multe niveluri. În primul rând, expresia BMP6 este suprimată; apoi, activitatea TMPRSS6, o protează care clivează hemojuvelinul co-receptorului BMP (Silvestri et al., 2008), este puternic crescută (Lakhal et al., 2011); și, în al treilea rând, histona deacetilaza3 (HDAC3) suprimă locusul hepcidinei (Pasricha et al., 2017). În condiții de deficiență de fier, reducerea producției de hepcidină este un mecanism de adaptare care facilitează absorbția fierului alimentar și farmacologic (Camaschella și Pagani, 2018).
Când anemia este severă, hipoxia coexistentă stimulează eritropoieza prin creșterea sintezei renale și eliberarea de EPO. Acest lucru duce la suprimarea transcrierii hepcidinei de către eritroferronă (ERFE), o genă țintă a EPO produsă de eritroblaste (Kautz et al., 2014), de către molecule (de exemplu, PDGF-BB) eliberate de alte țesuturi (Sonnweber et al., 2014) și, probabil, de către componentele solubile ale receptorilor de transferrină (TFR), sTFR1 (Beguin, 2003) și sTFR2 (Pagani et al., 2015). Scopul final este de a furniza suficient fier pentru nevoile unei eritropoieze extinse.
Anemii cu niveluri anormale de hepcidină
Anemiile pot fi clasificate pe baza nivelurilor de hepcidină în anemii cu hepcidină ridicată și scăzută. Este intuitiv faptul că nivelurile persistent ridicate de hepcidină, prin blocarea absorbției fierului, determină anemie feriprivă din cauza scăderii aportului de fier pentru eritropoieză. În schimb, eritropoieza ineficientă caracterizează așa-numitele anemii de încărcare cu fier, care au niveluri scăzute de hepcidină și supraîncărcare cu fier. Aceste două grupuri de anemii sunt rezultatul unor mecanisme fiziopatologice opuse (figura 1). În primul grup, anemia se datorează efectului inhibitor exercitat de hepcidină asupra absorbției și reciclării fierului care duce la deficit de fier sistemic; în cel de-al doilea grup, anemia se datorează suprimării hepcidinei printr-o eritropoieză anormală extinsă (Camaschella și Nai, 2016).
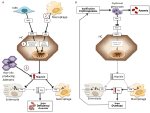
Figura 1. Reprezentarea schematică a mecanismelor anemiilor cu hepcidină ridicată (panoul din stânga) și scăzută (panoul din dreapta). Panoul (A). Patogeneza moleculară a anemiei asociate cu niveluri ridicate de hepcidină. LSEC, celule endoteliale sinusoidale hepatice producătoare de proteine morfogenetice osoase (BMPs); BMPRs, receptori BMP; IL6, interleukina 6; HC, hepatocite; HAMP, gena hepcidinei. Fe, fier; FPN, feroportină; 1, IRIDA; 2, anemie de inflamație; 3, adenom producător de hepcidină. Panoul (B). Patogenia moleculară a variației hepcidinei în anemiile datorate eritropoiezei ineficiente. ERFE, BMP-uri sechestratoare de eritroferrone. Alte mecanisme care inhibă hepcidina în acest tip de anemie, precum scăderea saturației transferinei și hipoxia, nu sunt prezentate. A se vedea textul pentru detalii.
Anemie asociată cu niveluri ridicate de hepcidină
Acest grup include două tulburări rare moștenite (anemie feriprivă refractară la fier și adenoame producătoare de hepcidină într-o eroare înnăscută a metabolismului glucozei) și o afecțiune comună dobândită: anemia de inflamație (tabelul 1).
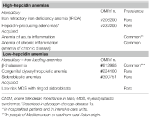
Tabelul 1. Anemii clasificate în funcție de nivelurile de hepcidină.
Anemie feriprivă refractară la fier
Anemia feriprivă refractară la fier (IRIDA) este o tulburare recesivă rară caracterizată prin anemie microcromă hipocromă microcitară, saturație scăzută a transferinei și niveluri de hepcidină inadecvat de normale/înalte. Este cauzată de mutații ale genei TMPRSS6 (Finberg et al., 2008), o genă care codifică o serin protează de tip II, matriptase-2 (Du et al., 2008). Mutațiile din TMPRSS6 sunt răspândite de-a lungul genei și pot afecta diferite domenii, în special domeniul catalitic (De Falco et al., 2014). Această protează transmembranară, puternic exprimată în ficat, inhibă transcrierea hepcidinei prin clivarea hemojuvelinului co-receptorului BMP de la suprafața celulară, atenuând astfel semnalizarea BMP și sinteza hepcidinei (Silvestri et al., 2008). Funcția TMPRSS6 este esențială în deficitul de fier pentru a permite mecanismul compensator de absorbție crescută a fierului.
IRIDA este prezentă încă de la naștere și, de obicei, este diagnosticată în copilărie. Comparativ cu deficitul de fier clasic, parametrii fierului sunt atipici și ridică suspiciunea bolii. Procentul de saturație a transferrinei este puternic redus (mai puțin de 10%), ca și în alte forme de deficit de fier; cu toate acestea, spre deosebire de deficitul de fier, nivelurile de feritină serică sunt normale/creșcute (Camaschella, 2013; De Falco et al., 2013). Acest lucru reflectă o acumulare crescută de feritină în macrofage, datorită nivelurilor ridicate de hepcidină care induc sechestrarea fierului din depozite.
Nici unul dintre testele propuse pentru diagnosticul IRIDA nu acoperă 100% din cazuri. Testul genetic identifică faptul că mutațiile TMPRSS6, care în unele cazuri (mutații non-sens, frame-shift și mutații de splicing), sunt în mod clar cauzale. În alte cazuri, ca în cazul mutațiilor missense neraportate anterior, sunt necesare studii funcționale pentru a demonstra cauzalitatea (Silvestri et al., 2013). Cu toate acestea, aceste teste sunt rareori disponibile. Nivelurile serice de hepcidină sunt de obicei crescute/normale, independent de deficitul de fier și în concordanță cu feritina ridicată/normală. Este important să se excludă inflamația prin dozarea concomitentă a proteinei C reactive.
S-a raportat că unii pacienți cu un fenotip de deficit de fier refractar au o singură alelă mutantă TMPRSS6; aici, dezbaterea este dacă aceștia ar trebui să fie considerați IRIDA sau nu. Se poate avea în vedere un spectru de afecțiuni care variază de la IRIDA severă clasică datorată mutațiilor TMPRSS6 homozigote sau heterozigote compuse până la o sensibilitate crescută la deficitul de fier conferită de mutații unice/modificări polimorfe. O abordare propusă pentru a prezice IRIDA clasică este normalizarea hepcidinei pe alți parametri de fier, ca raporturi saturație de transferrină (Tsat)/log hepcidină sau Tsat/log feritină (Donker et al., 2016). Potrivit altor autori, majoritatea pacienților cu un fenotip IRIDA sever au mutații TMPRSS6 bialelice și, atunci când nu sunt identificate, cea de-a doua alelă poate fi ascunsă din punct de vedere genetic (Heeney et al., 2018). În termeni generali, subiecții cu o singură alelă au un fenotip mai blând decât cei cu două mutații și răspund mai bine la tratamentul cu fier (Donker et al., 2016). În mod interesant, s-a demonstrat că mai multe SNP-uri TMPRSS6 oferă susceptibilitate la deficiența de fier la unele populații (An et al., 2012) și la donatorii de sânge (Sorensen et al., 2019).
A fost raportată o moștenire digenă la o fetiță în vârstă de 5 ani la care s-a constatat inițial că are un genotip IRIDA atipic cu o mutație cauzală TMPRSS6 (I212T) și o mutație silențioasă (R271Q) (De Falco et al., 2014). Ulterior, ea a fost diagnosticată cu Fibrodysplasia ossificans progressiva (FOP), o tulburare dominantă rară cu formare ectopică de os în țesuturile moi din cauza genei mutante a receptorului BMP de tip I ACVR1, care codifică ALK2 (Shore et al., 2006). Alela patologică ALK2R258S este constitutiv activă, deoarece mutația afectează domeniul bogat în glicină-serină al genei și face ca calea BMP/SMAD să fie hiperactivă, fiind incapabilă să se lege de inhibitorul său specific FKBP12 (Pagani et al., 2017).
Acest caz rar este deosebit de ilustrativ. În primul rând, deoarece domeniul bogat în glicină-serină ALK2 interacționează cu FKBP12, iar mutația destabilizează legătura, a dezvăluit un rol nesuspectat anterior pentru FKBP12 ca modulator al ALK2 și hepcidinei hepatice (Colucci et al., 2017). În al doilea rând, a dus la identificarea unei legături între activarea receptorilor BMP de tip I din os și ficat. În al treilea rând, cazul întărește relevanța TMPRSS6 intact în controlul semnalizării hepatice BMP/SMAD, deoarece nu a fost identificat niciun IRIDA în rândul altor pacienți FOP cu aceeași mutație ACVR1 și TMPRSS6 presupus normal (Pagani et al., 2017). În cele din urmă, acest caz este în concordanță cu conceptul că haploinsuficiența TMPRSS6 nu poate cauza IRIDA clasic.
Tratamentul optim al IRIDA este nedefinit. Fierul oral este ineficient, deoarece nu este absorbit. Adăugarea de vitamina C permite un răspuns sporadic. Fierul intravenos induce un răspuns parțial, de obicei într-un ritm mai lent în comparație cu pacienții cu deficit de fier dobândit. EPO este ineficientă în cazurile clasice (De Falco et al., 2013; Heeney și Finberg, 2014).
Anemia adenoamelor producătoare de hepcidină
Este o afecțiune extrem de rară la pacienții adulți afectați de boala de stocare a glicogenului 1a, o afecțiune recesivă datorată deficitului de gluco-6 fosfatază, care catalizează o reacție implicată atât în glicogenoliză, cât și în gluconeogeneză. Un simptom periculos comun al bolii este hipoglicemia. Tratamentul actual duce la o supraviețuire prelungită a copiilor afectați până la vârsta adultă, cu apariția mai multor complicații, cum ar fi anemia și adenoamele hepatice. Anemia este microcitară și hipocromă, deficitară în fier și refractară la tratamentul oral cu fier. Anemia a revenit după rezecția chirurgicală a adenomului. Țesutul adenomului a fost găsit pozitiv pentru ARNm de hepcidină, în timp ce țesutul normal din jur a prezentat suprimarea hepcidinei, așa cum era de așteptat din cauza producției ectopice necontrolate de hepcidină (Weinstein et al., 2002). Caracteristicile hematologice ale pacienților se aseamănă cu cele ale IRIDA, deoarece au în comun niveluri ridicate de hepcidină ca mecanism comun de anemie.
Anemia de inflamație
Anemia de inflamație (AI), cunoscută anterior ca anemia bolilor cronice, este o anemie moderată normocromă-normocitară care se dezvoltă în condiții de inflamație sistemică și activare imunitară. Apare în mai multe afecțiuni comune, inclusiv infecții cronice, boli autoimune, cancer avansat, boli cronice de rinichi, insuficiență cardiacă congestivă, boală pulmonară obstructivă cronică, anemie a vârstnicilor (cel puțin parțial) și boala grefei împotriva gazdei. IA este una dintre cele mai comune anemii la nivel mondial și cea mai frecventă anemie la pacienții spitalizați. Inflamația acută contribuie la severitatea anemiei în unitățile de terapie intensivă. Mecanismele moleculare care stau la baza IA sunt multiple și complexe. Supraproducția de citokine, cum ar fi IL1-β, TNF-α și IL-6 de către macrofage și INF-γ de către limfocite, frânează producția de EPO, afectează răspunsul la eritropoieză, crește nivelul de hepcidină și poate activa eritrofagocitoza, în special în formele acute (Weiss și Goodnough, 2005; Ganz, 2019).
Hepcidina este activată de IL-6 prin intermediul receptorului IL-6 (IL-6R) și a semnalizării JAK2-STAT3. Activarea completă a hepcidinei necesită o cale BMP-SMAD activă, deoarece inactivarea semnalizării BMP scade hepcidina în modelele animale de inflamație (Theurl et al., 2011). Dereglarea homeostaziei fierului sistemic determină sechestrarea fierului de către macrofage și reducerea absorbției și reciclării, ceea ce duce la o saturație scăzută a transferrinei și la restricția de fier a eritropoiezei și a altor țesuturi.
Tratamentul tradițional al IA se bazează pe reversibilitatea/controlul bolii de bază, ori de câte ori este posibil. Dacă boala este netratabilă și anemia este ușoară, este necesară o evaluare atentă a raportului riscuri-beneficii pentru a evita efectele secundare ale oricărui tratament. Tratamentele bazate pe fiziopatologie se limitează la compuși asemănători eritropoietinei și la fier. Utilizarea agenților de stimulare a eritropoiezei (ESA) suprimă hepcidina prin inducerea expansiunii eritropoiezei. Această abordare este utilizată pe scară largă la pacienții cu boală cronică de rinichi, sindroame mielodisplazice cu risc scăzut și cancer care fac chimioterapie. Cu toate acestea, este necesar un control clinic atent, deoarece dozele mari au efecte secundare cardiovasculare. Administrarea de fier intravenos poate ameliora restricția de fier, cauzată de expansiunea eritropoiezei dependentă de ESA. Fierul oral este de obicei ineficient, deoarece nivelurile ridicate de hepcidină contracarează absorbția sa intestinală. Inhibitorii de prolil hidroxilază (stabilizatori ai factorului inductibil de hipoxie, HIF) sunt experimentali în boala cronică de rinichi, cu scopul de a crește EPO endogenă. Tratamentul cronic cu transfuzii de globule roșii nu este recomandat din cauza efectului tranzitoriu și a reacțiilor adverse; acesta se limitează la anemiile refractare severe (Camaschella, 2019; Weiss et al., 2019).
Anemii asociate cu niveluri scăzute de hepcidină
Eropoieza ineficientă și nivelurile scăzute sau inadecvat de normale ale hepcidinei, cu supraîncărcarea cu fier consecutivă, sunt caracteristici tipice ale „anemiilor cu încărcare cu fier”.” Prototipul este β-talasemia, o boală genetică recesivă datorată unor mutații ale genei β-globinei care provoacă anemie și producerea în exces a lanțului α-globinei. Aceasta din urmă precipită sub formă de hemicromi în măduva osoasă, deteriorând precursorii eritroizi în curs de maturare și ducând la o eritropoieză ineficientă. Acest lucru se întâmplă în cazul talasemiei netransfuzionale dependente sau al talasemiei intermedia, a cărei eritropoieză se caracterizează prin prevalența celulelor imature care eliberează eritroferronă pentru a inhiba expresia hepcidinei hepatice. Nivelurile de hepcidină sunt de obicei mai mari în talasemia dependentă de transfuzii, în care eritropoieza ineficientă endogenă este cel puțin parțial suprimată de transfuzii (Camaschella și Nai, 2016).
Supresia hepcidinei este mediată de creșterea cantității de citokină eritroferronă (ERFE), un membru al familiei TNF-α codificat de gena ERFE, sintetizată de eritroblaste la stimularea cu EPO (Kautz et al., 2014). ERFE este eliberată în circulație și sechestrează BMP-urile, în special BMP6 (Arezes et al., 2018), atenuând semnalizarea hepcidinei ca răspuns la fier. În plus, are loc o suprimare epigenetică la nivelul locusului hepcidinei de către histone deacetilaza HDAC3 (Pasricha et al., 2017). Atunci când anemia provoacă hipoxie, alți mediatori, cum ar fi PDGF-BB (Sonnweber et al., 2014), care este eliberat de diferite tipuri de celule, suprimă hepcidina.
Nivelurile de hepcidină sunt diminuate printr-un mecanism special în mielodisplazia cu risc scăzut cu sideroblaste inelate, o afecțiune clonală datorată mutațiilor genei spliceosomului SF3B1. Fierul se acumulează în mitocondrii, ceea ce duce la eritropoieză ineficientă și supraîncărcare sistemică cu fier. O proteină ERFE alungită, splicată anormal, este mai puternică decât ERFE de tip sălbatic în suprimarea hepcidinei (Bondu et al., 2019) și provocând o încărcare cu fier independentă de transfuzii.
Terapii țintite pentru anemiile legate de hepcidină
Identificarea mecanismelor moleculare responsabile de anemiile discutate anterior a stimulat cercetarea în dezvoltarea de terapii țintite pentru a înlocui tratamentul simptomatic actual (Sebastiani et al., 2016; Crielaard et al., 2017). Abordările diferă în funcție de tipul de anemie și de obiectivul de scădere sau de creștere a nivelului de hepcidină sau a efectelor acestora (Tabelul 2).
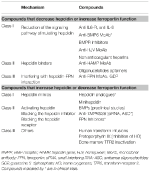
Tabel 2. Terapii experimentale care vizează axa hepcidină-ferroportină.
Terapii experimentale pentru scăderea nivelurilor de hepcidină/creșterea funcției feroportinei
Cu excepția tumorilor producătoare de hepcidină, care trebuie îndepărtate chirurgical, compușii care antagonizează hepcidina sau efectele acesteia pot fi utili în toate anemiile caracterizate prin niveluri ridicate de hepcidină. Principala lor aplicație ar fi în bolile inflamatorii cronice, pentru a inversa hipoferremia și anemia. Mai multe terapii experimentale care vizează manipularea căii hepcidinei și a funcției acesteia au fost investigate în studii preclinice. Antagoniștii hepcidinei sunt inhibitori ai sintezei/regulatorilor hepcidinei (Ganz, 2019), lianți ai hepcidinei care blochează funcția acesteia și compuși care interferează cu interacțiunea hepcidină-ferroportină (tabelul 2). Unii compuși se află în studii clinice, în special în boala cronică de rinichi (Sheetz et al., 2019). În IRIDA, manipularea căii hepcidinei a fost propusă în studiile preclinice cu utilizarea de MoAb anti-HJV (Kovac et al., 2016).
Terapii experimentale pentru creșterea nivelurilor de hepcidină/diminuarea funcției feroportinei
Creșterea nivelurilor de hepcidină poate nu numai să reducă supraîncărcarea cu fier, ci și să controleze parțial eritropoieza ineficientă în anemiile cu încărcare cu fier. β-talasemia este cea mai studiată dintre aceste afecțiuni (Casu et al., 2018; Gupta et al., 2018). Medicamentele propuse sunt analogi ai hepcidinei (unii în studii clinice), modulatori ai hepcidinei, în special inhibitori ai TMPRSS6, sau compuși care interferează cu interacțiunea hepcidină-ferroportină scăzând exportul de fier (tabelul 2).
În timp ce compușii care cresc hepcidina reduc eritropoieza ineficientă din cauza cercului vicios dintre eritropoieza ineficientă și încărcarea cu fier (Camaschella și Nai, 2016), medicamentele care favorizează maturarea precursorilor eritroizi, precum capcana ligandului receptorului IIB al activinei, luspatercept, nu numai că îmbunătățesc anemia, dar ameliorează și homeostazia fierului prin reducerea inhibiției hepcidinei (Piga et al., 2019).
Se speră că unele abordări țintite aflate acum în studii clinice vor avea ca rezultat tratamente noi pentru o varietate de anemii.
Concluzie
Progresele spectaculoase în înțelegerea reglementării metabolismului fierului și a hepcidinei au permis o mai bună înțelegere a controlului eritropoiezei, deoarece, împreună cu eritropoietina, fierul este un factor fundamental pentru maturarea celulelor eritroide. Afecțiunile care duc la anemie pot fi asociate cu niveluri ridicate și scăzute de hepcidină. În ambele cazuri, dereglarea contrastantă a hepcidinei poate ameliora/corecta anemia în modele preclinice, oferind noi instrumente care sunt deja sau vor fi în curând explorate clinic pentru tratamentul anemiilor specifice.
Contribuții ale autorilor
AP a redactat lucrarea. CC a elaborat versiunea finală. AN și LS au contribuit la redactarea și la revizuirea critică a manuscrisului. Toți autorii au aprobat versiunea finală.
Finanțare
Acest articol a fost susținut parțial de un grant de cercetare avansată EHA în 2018 pentru AP. Cariplo Foundation Young Investigator Grant nr. 2017-0916 către AN.
Conflict de interese
CC este consultant al Vifor Pharma, Celgene și Novartis.
Ceilalți autori declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
An, P., Wu, Q., Wang, H., Guan, Y., Mu, M., Liao, Y., et al. (2012). Variantele TMPRSS6, dar nu TF, TFR2 sau BMP2, sunt asociate cu un risc crescut de anemie cu deficit de fier. Hum. Mol. Genet. 21, 2124-2131. doi: 10.1093/hmg/dds028
PubMed Abstract | CrossRef Full Text | Google Scholar
Andriopoulos, B. Jr., Corradini, E., Xia, Y., Faasse, S. A., Chen, S., Grgurevic, L., et al. (2009). BMP6 este un regulator endogen cheie al expresiei hepcidinei și al metabolismului fierului. Nat. Genet. 41, 482-487. doi: 10.1038/ng.335
PubMed Abstract |Ref Full Text | Google Scholar
Arezes, J., Foy, N., McHugh, K., Sawant, A., Quinkert, D., Terraube, V., et al. (2018). Erythroferrone inhibă inducerea hepcidinei de către BMP6. Blood 132, 1473-1477. doi: 10.1182/blood-2018-06-857995
PubMed Abstract | CrossRef Full Text | Google Scholar
Aschemeyer, S., Qiao, B., Stefanova, D., Valore, E. V., Sek, A. C., Ruwe, T. A., et al. (2018). Analiza structură-funcție a ferroportinei definește locul de legare și un mecanism alternativ de acțiune a hepcidinei. Sânge. 131, 899-910. doi: 10.1182/blood-2017-05-786590
CrossRef Full Text | Google Scholar
Babitt, J. L., Huang, F. W., Wrighting, D. M., Xia, Y., Sidis, Y., Samad, T. A., et al. (2006). Semnalizarea proteinei morfogenetice osoase de către hemojuvelin reglează expresia hepcidinei. Nat. Genet. 38, 531-539. doi: 10.1038/ng1777
PubMed Abstract | Text integral | Google Scholar
Beguin, Y. (2003). Receptor solubil de transferrină pentru evaluarea eritropoiezei și a statusului de fier. Clin. Chim. Acta 329, 9-22. doi: 10.1016/S0009-8981(03)00005-6
PubMed Abstract | Refef Full Text | Google Scholar
Bondu, S., Alary, A. S., Lefevre, C., Houy, A., Jung, G., Lefebvre, T., et al. (2019). O variantă a eritroferronei perturbă homeostazia fierului în sindromul mielodisplastic SF3B1-mutat. Sci. Transl. Med. 11:pii:eaav5467. doi: 10.1126/scitranslmed.aav5467
PubMed Abstract | CrossRef Full Text | Google Scholar
Camaschella, C. (2013). Cum gestionez pacienții cu anemie microcitară atipică. Br. J. Haematol. 160, 12-24. doi: 10.1111/bjh.12081
PubMed Abstract |Ref Full Text | Google Scholar
Camaschella, C. (2019). Deficiența de fier. Blood 133, 30-39. doi: 10.1182/blood-2018-05-05-815944
PubMed Abstract | CrossRef Full Text | Google Scholar
Camaschella, C., și Nai, A. (2016). Eritropoieza ineficientă și reglarea statusului de fier în anemiile cu încărcare cu fier. Br. J. Haematol. 172, 512-523. doi: 10.1111/bjh.13820
PubMed Abstract | Full CrossRef Text | Google Scholar
Camaschella, C., și Pagani, A. (2018). Progrese în înțelegerea metabolismului fierului și a crosstalk-ului său cu eritropoieza. Br. J. Haematol. 182, 481-494. doi: 10.1111/bjh.15403
PubMed Abstract | Full CrossRef Text | Google Scholar
Canali, S., Wang, C. Y. Y., Zumbrennen-Bullough, K. B., Bayer, A., și Babitt, J. L. (2017). Proteina morfogenetică osoasă 2 controlează homeostazia fierului la șoareci independent de Bmp6. Am. J. Hematol. 92, 1204-1213. doi: 10.1002/ajh.24888
PubMed Abstract | Full Text | Google Scholar
Casu, C., Nemeth, E., și Rivella, S. (2018). Agoniștii de hepcidină ca instrumente terapeutice. Blood 131, 1790-1794. doi: 10.1182/blood-2017-11-737411
PubMed Abstract | CrossRef Full Text | Google Scholar
Colucci, S., Pagani, A., Pettinato, M., Artuso, I., Nai, A., Camaschella, C., et al. (2017). Imunofilina FKBP12 inhibă expresia hepcidinei prin legarea receptorului BMP de tip I ALK2 în hepatocite. Blood 130, 2111-2120. doi: 10.1182/blood-2017-04-780692
PubMed Abstract | Full CrossRef Text | Google Scholar
Crielaard, B. J., Lammers, T., și Rivella, S. (2017). Direcționarea metabolismului fierului în descoperirea și livrarea de medicamente. Nat. Rev. Drug Discov. 16, 400-423. doi: 10.1038/nrd.2016.248
PubMed Abstract | Text integral | Google Scholar
De Falco, L., Sanchez, M., Silvestri, L., Kannengiesser, C., Muckenthaler, M. U., Iolascon, A., et al. (2013). Anemia feriprivă refractară la fier. Haematologica 98, 845-853. doi: 10.3324/haematol.2012.075515
PubMed Abstract | CrossRef Full Text | Google Scholar
De Falco, L., Silvestri, L., Kannengiesser, C., Moran, E., Oudin, C., Rausa, M., et al. (2014). Impactul funcțional și clinic al noilor variante TMPRSS6 la pacienții cu anemie feriprivă refractară la fier și studii de genotip-fenotip. Hum. Mutat. 35, 1321-1329. doi: 10.1002/humu.22632
PubMed Abstract | CrossRefect Full Text | Google Scholar
Donker, A. E., Schaap, C. C., Novotny, V. M., Smeets, R., Peters, T. M., van den Heuvel, B. L., et al. (2016). Anemia feriprivă refractară la fier: o boală eterogenă care nu este întotdeauna refractară la fier. Am. J. Hematol. 91, E482-E490. doi: 10.1002/ajh.24561
PubMed Abstract | Full Text | Google Scholar
Du, X., She, E., Gelbart, T., Truksa, J., Lee, P., Xia, Y., et al. (2008). Serina protează TMPRSS6 este necesară pentru a detecta deficiența de fier. Science 320, 1088-1092. doi: 10.1126/science.1157121
PubMed Abstract | CrossRef Full Text | Google Scholar
Finberg, K. E., Heeney, M. M., Campagna, D. R., Aydinok, Y., Pearson, H. A., Hartman, K. R., et al. (2008). Mutațiile în TMPRSS6 cauzează anemia feriprivă refractară la fier (IRIDA). Nat. Genet. 40, 569-571. doi: 10.1038/ng.130
PubMed Abstract |Refef Full Text | Google Scholar
Ganz, T. (2019). Anemia de inflamație. N. Engl. J. Med. 381, 1148-1157. doi: 10.1056/NEJMra1804281
PubMed Abstract | Text integral | Google Scholar
Gupta, R., Musallam, K. M., Taher, A. T., și Rivella, S. (2018). Eritropoieza ineficientă: anemie și supraîncărcare cu fier. Hematol. Oncol. Clin. North Am. 32, 213-221. doi: 10.1016/j.hoc.2017.11.009
PubMed Abstract | CrossRef Full Text | Google Scholar
Heeney, M. M., și Finberg, K. E. (2014). Anemia feriprivă refractară la fier (IRIDA). Hematol. Oncol. Clin. North Am. 28, 637-652. doi: 10.1016/j.hoc.2014.04.009
PubMed Abstract | Cross Full Text | Google Scholar
Heeney, M. M., Guo, D., De Falco, L., Campagna, D. R., Olbina, G., Kao, P. P., et al. (2018). Normalizarea hepcidinei prezice starea de mutație TMPRSS6 la pacienții cu deficiență cronică de fier. Blood 132, 448-452. doi: 10.1182/blood-2017-03-773028
PubMed Abstract | CrossRef Full Text | Google Scholar
Kautz, L., Jung, G., Valore, E. V., Rivella, S., Nemeth, E., și Ganz, T. (2014). Identificarea eritroferronei ca un regulator eritroid al metabolismului fierului. Nat. Genet. 46, 678-684. doi: 10.1038/ng.2996
PubMed Abstract | Refef Full Text | Google Scholar
Koch, P. S., Olsavszky, V., Ulbrich, F., Sticht, C., Demory, A., Leibing, T., et al. (2017). Semnalizarea angiocrină Bmp2 în ficatul murin controlează homeostazia normală a fierului. Blood 129, 415-419. doi: 10.1182/blood-2016-07-729822
PubMed Abstract | CrossRef Full Text | Google Scholar
Kovac, S., Boser, P., Cui, Y., Ferring-Appel, D., Casarrubea, D., Huang, L., et al. (2016). Anticorpul antihemojuvelină corectează anemia cauzată de niveluri inadecvate de hepcidină. Haematologica 101, e173-e176. doi: 10.3324/haematol.2015.140772
PubMed Abstract | CrossRef Full Text | Google Scholar
Lakhal, S., Schodel, J., Townsend, A. R., Pugh, C. W., Ratcliffe, P. J., și Mole, D. R. (2011). Reglarea serin-proteinazei serice transmembranare de tip II TMPRSS6 de către factorii induși de hipoxie: o nouă legătură între semnalizarea hipoxiei și homeostazia fierului. J. Biol. Chem. 286, 4090-4097. doi: 10.1074/jbc.M110.173096
PubMed Abstract | CrossRef Full Text | Google Scholar
Meynard, D., Kautz, L., Darnaud, V., Canonne-Hergaux, F., Coppin, H., și Roth, M. P. (2009). Lipsa proteinei morfogenetice osoase BMP6 induce o supraîncărcare masivă de fier. Nat. Genet. 41, 478-481. doi: 10.1038/ng.320
PubMed Abstract |Ref Full Text | Google Scholar
Muckenthaler, M. U., Rivella, S., Hentze, M. W., și Galy, B. (2017). Un covor roșu pentru metabolismul fierului. Cell 168, 344-361. doi: 10.1016/j.cell.2016.12.034
PubMed Abstract | Full CrossRef Text | Google Scholar
Nemeth, E., Tuttle, M. S., Powelson, J., Vaughn, M. B., Donovan, A., Ward, D. M., et al. (2004). Hepcidina reglează efluxul celular de fier prin legarea la feroportină și prin inducerea internalizării acesteia. Science 306, 2090-2093. doi: 10.1126/science.1104742
PubMed Abstract | CrossRef Full Text | Google Scholar
Pagani, A., Colucci, S., Bocciardi, R., Bertamino, M., Dufour, C., Ravazzolo, R., et al. (2017). O nouă formă de IRIDA datorată mutațiilor heterozigote combinate ale TMPRSS6 și ACVR1A care codifică receptorul BMP ALK2. Blood 129, 3392-3395. doi: 10.1182/blood-2017-03-773481
PubMed Abstract | Publicitate integrală a textului | Google Scholar
Pagani, A., Vieillevoye, M., Nai, A., Rausa, M., Ladli, M., Lacombe, C., et al. (2015). Reglarea receptorului 2 al transferrinei de suprafață celulară prin scindarea dependentă de fier și eliberarea unei forme solubile. Haematologica 100, 458-465. doi: 10.3324/haematol.2014.118521
PubMed Abstract | CrossRef Full Text | Google Scholar
Pasricha, S. R., Lim, P. J., Duarte, T. L., Casu, C., Oosterhuis, D., Mleczko-Sanecka, K., et al. (2017). Hepcidina este reglată de acetilarea histonică asociată promotorului și de HDAC3. Nat. Commun. 8:403. doi: 10.1038/s41467-017-00500-z
PubMed Abstract | Full CrossRef Text | Google Scholar
Piga, A., Perrotta, S., Gamberini, M. R., Voskaridou, E., Melpignano, A., Filosa, A., et al. (2019). Luspatercept îmbunătățește nivelurile de hemoglobină și necesarul de transfuzii de sânge într-un studiu de pacienți cu beta-talasemie. Blood 133, 1279-1289. doi: 10.1182/blood-2018-10-879247
PubMed Abstract | Ref Full Text | Google Scholar
Rausa, M., Pagani, A., Nai, A., Campanella, A., Gilberti, M. E., Apostoli, P., et al. (2015). Expresia Bmp6 în celulele non-parenchimatoase ale ficatului murin: un mecanism de control al activității lor de exportator de fier ridicat și de protecție a hepatocitelor împotriva supraîncărcării cu fier? PLoS One 10:e0122696. doi: 10.1371/journal.pone.0122696
PubMed Abstract | Full CrossRef Text | Google Scholar
Sebastiani, G., Wilkinson, N., și Pantopoulos, K. (2016). Direcționarea farmacologică a axei hepcidină/ferroportină. Front. Pharmacol. 7:160. doi: 10.3389/fphar.2016.00160
PubMed Abstract | Full CrossRef Text | Google Scholar
Sheetz, M., Barrington, P., Callies, S., Berg, P. H., McColm, J., Marbury, T., et al. (2019). Direcționarea căii hepcidină-ferroportină în anemia bolii renale cronice. Br. J. Clin. Pharmacol. 85, 935-948. doi: 10.1111/bcp.13877
PubMed Abstract | Text integral | Google Scholar
Shore, E. M., Xu, M., Feldman, G. J., Fenstermacher, D. A., Cho, T. J., Choi, I. H., et al. (2006). O mutație recurentă în receptorul BMP de tip I ACVR1 cauzează fibrodisplazia osifică progresivă moștenită și sporadică. Nat. Genet. 38, 525-527. doi: 10.1038/ng1783
PubMed Abstract | CrossRef Full Text | Google Scholar
Silvestri, L., Pagani, A., Nai, A., De Domenico, I., Kaplan, J., și Camaschella, C. (2008). Serina protează matriptase-2 (TMPRSS6) inhibă activarea hepcidinei prin clivarea hemojuvelinei membranare. Cell Metab. 8, 502-511. doi: 10.1016/j.cmet.2008.09.012
PubMed Abstract | CrossRef Full Text | Google Scholar
Silvestri, L., Rausa, M., Pagani, A., Nai, A., și Camaschella, C. (2013). Cum să evaluăm cauzalitatea mutațiilor TMPRSS6? Hum. Mutat. 34, 1043-1045. doi: 10.1002/humu.22321
PubMed Abstract | Full CrossRef Text | Google Scholar
Sonnweber, T., Nachbaur, D., Schroll, A., Nairz, M., Seifert, M., Demetz, E., et al. (2014). Downregulation indusă de hipoxie a hepcidinei este mediată de factorul de creștere BB derivat din trombocite. Gut 63, 1951-1959. doi: 10.1136/gutjnl-2013-305317
PubMed Abstract | Refef Full Text | Google Scholar
Sorensen, E., Rigas, A. S., Didriksen, M., Burgdorf, K. S., Thorner, L. W., Pedersen, O. B., et al. (2019). Factori genetici care influențează nivelul hemoglobinei la 15.567 de donatori de sânge: rezultate din studiul danez al donatorilor de sânge. Transfusion 59, 226-231. doi: 10.1111/trf.15075
CrossRef Full Text | Google Scholar
Theurl, I., Schroll, A., Sonnweber, T., Nairz, M., Theurl, M., Willenbacher, W., et al. (2011). Inhibarea farmacologică a expresiei hepcidinei inversează anemia de inflamație cronică la șobolani. Blood 118, 4977-4984. doi: 10.1182/blood-2011-03-345066
PubMed Abstract | CrossRef Full Text | Google Scholar
Weinstein, D. A., Roy, C. N., Fleming, M. D., Loda, M. F., Wolfsdorf, J. I., și Andrews, N. C. (2002). Expresia nepotrivită a hepcidinei este asociată cu anemia refractară la fier: implicații pentru anemia bolilor cronice. Blood 100, 3776-3781. doi: 10.1182/blood-2002-04-1260
PubMed Abstract | CrossRef Full Text | Google Scholar
Weiss, G., Ganz, T., and Goodnough, L. T. (2019). Anemia de inflamație. Blood 133, 40-50. doi: 10.1182/blood-2018-06-856500
PubMed Abstract | CrossRef Full Text | Google Scholar
Weiss, G. și Goodnough, L. T. (2005). Anemia de boală cronică. N. Engl. J. Med. 352, 1011-1023. doi: 10.1056/NEJMra041809
PubMed Abstract | Full CrossRef Full Text | Google Scholar
.
Leave a Reply