Frontiers in Celland Developmental Biology
Background
De basiseenheid van het Golgi-apparaat wordt gewoonlijk beschouwd als een stapel cisternae die sterk gepolariseerd is, met de cis-face ontvangen lading van de ER en de trans-face van de stapel, de TGN, geassocieerd met het sorteren van lading voor post-Golgi export (Boncompain en Perez, 2013). De organisatie van deze mini-stapels in de cel varieert echter tussen verschillende organismen. In planten en ongewervelden zijn individuele Golgi stapels onafhankelijk verspreid over het cytoplasma, terwijl in de meeste gewervelde cellen tijdens de interfase, individuele Golgi stapels zijn samengesmolten in een compacte lintstructuur gelegen in de nabijheid van de MTOC (Wei en Seemann, 2017). De structuur van het Golgi-lint in zoogdiercellen wordt het best onthuld door elektronenmicroscopie (Rambourg en Clermont, 1997); optische microscopie met hoge resolutie met behulp van cis- en trans-markers kan de lintorganisatie ook detecteren (Gosavi et al., 2018). Een belangrijke vraag die in het veld wordt onderzocht is de relevantie van de Golgi lintstructuur en de functionele verschillen van het Golgi lint in vergelijking met “geïsoleerde Golgi mini-stacks” of andere toestanden van Golgi architectuur. Met andere woorden, wat is het evolutionaire voordeel van de meer complexe lintmorfologie van het Golgi-apparaat in gewervelde cellen en welke functies kunnen worden gereguleerd door een overgang naar een niet-ribbon structuur?
Studies van de laatste jaren tonen aan dat Golgi-membranen een platform bieden voor de regulatie van een reeks cellulaire processen, waaronder celpolarisatie (Kupfer et al., 1983), gerichte migratie (Millarte en Farhan, 2012), stress (Sasaki en Yoshida, 2015), DNA-reparatie (Farber-Katz et al., 2014), mitose (Rabouille en Kondylis, 2007), metabolisme (Abdel Rahman et al., 2015), pro-inflammatoire reacties (Chen en Chen, 2018) en autofagie (Yamamoto et al., 2012; Lamb et al., 2013). In zoogdiercellen zijn er nu inderdaad aanzienlijke aanwijzingen dat de Golgi, net als andere intracellulaire organellen, kan fungeren als een celsensor (Farhan en Rabouille, 2011; Mayinger, 2011; Millarte en Farhan, 2012; Sasaki en Yoshida, 2015; Luini en Parashuraman, 2016; Gosavi en Gleeson, 2017; Makhoul et al., 2018). Het wordt ook duidelijk dat de precieze morfologie van de Golgi relevant is voor de regulatie van een aantal van deze celprocessen (Makhoul et al., 2018). De associatie van Golgi morfologie met signalering kwam voort uit een genoom breed kinoom en fosfatoom scherm dat een groot cohort van kinasen en fosfatasen (20% van het totaal in het genoom) identificeerde die de morfologie van de Golgi beïnvloedde (Chia et al., 2012). De veranderingen in Golgi morfologie omvatten ofwel fragmentatie van de Golgi (verlies van Golgi lint) of de vorming van een zeer compacte en gecondenseerde Golgi in de perinucleaire locatie. De relevantie van actine in de regulatie van de Golgi-morfologie werd in deze studie benadrukt door de identificatie van een aantal kinasen, bijvoorbeeld ROCK1 en PAK1, die de actinedynamiek reguleren en de Golgi-structuur moduleren (Chia et al., 2012). Andere genoombrede analyses hebben ook de waarschijnlijkheid benadrukt dat de Golgi een grote verscheidenheid aan signalen kan ontvangen en doorgeven die niet alleen membraantransportroutes kunnen beïnvloeden, maar ook andere processen, apoptose, mitose, autofagie en stressreacties (Farhan et al., 2010; Millarte et al., 2015).
De Golgi-lintstructuur is zeer dynamisch en kan een zeer snelle hermodellering ondergaan tijdens een reeks verschillende omstandigheden. Bijvoorbeeld, tijdens mitose de demontage van het Golgi-lint is een vroege gebeurtenis in G2/M overgang en speelt een belangrijke rol als een celcyclus checkpoint in het bevorderen van mitotische entry (Wei en Seemann, 2010; Corda et al., 2012). De regulatie van Golgi dynamiek wordt gemedieerd door interacties tussen moleculaire steigers gelegen op de Golgi membraan en het cytoskelet. MT dynamiek kan de locatie van het Golgi-lint bij het centrosoom en de herpositionering van de Golgi reguleren om gepolariseerde trafficking en gerichte secretie te vergemakkelijken (Millarte en Farhan, 2012; Sanders en Kaverina, 2015). Bovendien kunnen membraancomponenten van de Golgi ook microtubuli kernen en stabiliseren bij zowel de cis- als trans-Golgi, en daarom is de Golgi zelf ook een MTOC (Efimov et al., 2007; Wu et al., 2016). Actine-gemedieerde processen dragen ook bij aan de vorm en functie van de Golgi en ten minste negen Golgi-gelokaliseerde moleculaire scaffolds zijn geïdentificeerd die interageren met het actine cytoskelet . Versterking van actinepolymerisatie bij de Golgi resulteert in verspreiding van het lint, terwijl remming van actinepolymerisatie met specifieke geneesmiddelen zoals latrunculine A resulteert in verdichting van de Golgi (Lazaro-Dieguez et al., 2006; Makhoul et al., 2018, 2019). Gezien deze regulerende netwerken die de Golgi-lintstructuur moduleren, achten wij het zeer aannemelijk dat het evenwicht tussen het Golgi-lint en Golgi-ministapels zowel de kwalitatieve als de kwantitatieve respons van signaalwegen kan bepalen. Het verband tussen Golgi morfologie en signaaltransductie heeft ook belangrijke vertakkingen in het begrijpen van de moleculaire basis van een aantal ziekten die geassocieerd worden met het verlies van het Golgi lint en het verschijnen van een verspreide Golgi. Zo is bijvoorbeeld aangetoond dat de overleving van sommige kankercellen geassocieerd is met een verspreide Golgi die het niveau van apoptose vermindert (Farber-Katz et al., 2014; Petrosyan, 2015).
Wat betekent Golgi-fragmentatie?
De term “Golgi-fragmentatie” wordt algemeen gebruikt om de morfologische status van een verspreide Golgi in zoogdiercellen te beschrijven, zoals gedetecteerd door optische microscopie gekleurd met Golgi-markers. Een verspreide Golgi wordt vaak waargenomen in experimentele systemen, bijvoorbeeld wanneer cellen worden behandeld met geneesmiddelen om het cytoskelet te verstoren, bijvoorbeeld nocodazol (Wei en Seemann, 2010), wanneer membraan flux wordt verstoord, of wanneer componenten van Golgi transport machines, of het cytoskelet interactie systeem, worden neergehaald, uitgeklopt, of overgeëxpresseerd (Zappa et al., 2018). In verschillende fysiologische toestanden, bijvoorbeeld stress (Serebrenik et al., 2018) en pathologische omstandigheden, met name kanker en neurodegeneratie, heeft de Golgi vaak de typische compacte juxtanucleaire locatie verloren en wordt waargenomen door confocale microscopie als verspreide structuren in het hele cytoplasma . Er zijn een toenemend aantal rapporten beschrijven Golgi fragmentatie onder fysiologische en pathologische omstandigheden (Figuur 1A). Echter, een probleem met het gebruik van de term “gefragmenteerd” is dat het impliceert dat de structurele integriteit van de Golgi verloren is gegaan en dat de morfologie in verband met de “gefragmenteerde” structuur vertegenwoordigt een uiteengevallen, abnormale of vernietigd organel. In een aantal gevallen is dit duidelijk misleidend, aangezien de afzonderlijke Golgistapels intact kunnen blijven en de klassieke functies van de organel, namelijk glycosylering en membraantransport, kunnen blijven vervullen. Inderdaad, verspreide Golgi mini-stapels komen voor in sommige gespecialiseerde cellen, zoals gedifferentieerde myoblasten (Lu et al., 2001), gedifferentieerde neuronen die individuele Golgi stapels of “buitenposten” langs dendrieten bevatten (Lasiecka en Winckler, 2011), maag pariëtale cellen (Gunn et al., 2011), en uroepitheliale cellen van de urineblaas (Kreft et al., 2010), zonder schijnbare tekortkomingen in membraantransport en glycosylatie. Integendeel, de verspreiding van het Golgi lint in verband met experimentele en pathologische omstandigheden kan een verschuiving van het dynamisch evenwicht tussen de compacte lint morfologie en de individuele Golgi mini-stapels weerspiegelen of kan resulteren in verstoring van de lint architectuur en de integriteit van de Golgi stapel. Dit is een relevante kwestie omdat de precieze morfologische status van de Golgi zeer waarschijnlijk van invloed zal zijn, in sommige gevallen op de efficiëntie van het transport en de glycosylering (Puthenveedu et al., 2006), en in andere gevallen op een verscheidenheid van signaleringsnetwerken, maar niet noodzakelijkerwijs op het transport of de glycosylering. Het is belangrijk om onderscheid te maken tussen intacte Golgi mini-stacks en verlies van de integriteit van de Golgi stacks bij het definiëren van wat wordt bedoeld met een “gefragmenteerde” Golgi. Hier bespreken we de verschillende Golgi morfologieën die zijn gedetecteerd en gekarakteriseerd in experimentele, fysiologische en pathologische setting.
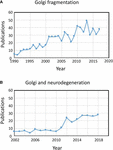
Figuur 1. Publicaties waarin fragmentatie van het Golgi-lint wordt geïdentificeerd. (A) Aantal publicaties per jaar met de term Golgi-fragmentatie in de titel of het uittreksel. (B) Aantal publicaties per jaar waarin de Golgi is onderzocht bij neurodegeneratieve ziekten. Gegevens zijn afkomstig van Alexandru Dan Corlan. Medline trend: geautomatiseerde jaarstatistieken van PubMed-resultaten voor elke query, 2004. Web bron op URL: http://dan.corlan.net/medline-trend.html. Accessed: 2019-04-29.
Rethinking the Terminology of Golgi Morphological States
De structuren van de Golgi-fragmenten verschillen afhankelijk van de aard van de pathway die betrokken is bij het verstoren of moduleren van het Golgi-lint. Het is belangrijk om een betere ultrastructurele karakterisering van de Golgi “fragmenten” na het verlies van het Golgi lint als de functionele uitkomst is waarschijnlijk zeer verschillend, afhankelijk van de precieze Golgi structuren. Afgezien van de voorwaarden die resulteren in een langgerekte Golgi lint, kunnen we identificeren uit de literatuur ten minste 4 verschillende scenario’s in verband met Golgi lint “fragmentatie.” Deze zijn afgebeeld in figuur 2 en als volgt beschreven:
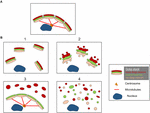
Figuur 2. Model met verschillende Golgi-morfologieën na “fragmentatie” van de Golgi-lintstructuur. (A) Intact Golgi lint structuur en (B) verschillende scenario’s met verlies van Golgi lint. (1) Een scenario waar intact Golgi mini-stacks zijn verspreid in het cytoplasma, (2) Een scenario waar de integriteit van de verspreide Golgi stapels is aangetast met verkorte cisternae, zwelling van cisternae en de toename van Golgi geassocieerde tubuli en blaasjes, (3) Een scenario waar er verspreiding van een Golgi compartiment. Hier wordt de TGN selectief verspreid over het cytoplasma terwijl de rest van de stack in een lintstructuur blijft; (4) Scenario waar er verlies is van lint en stapels met Golgi-membranen verspreid overwegend als tubuli en vesikels. Nummers verwijzen naar de classificatie van de Golgi morfologieën gegeven in de tekst.
(1) Omzetting van Golgi lint naar Golgi stapels. Hier gaat het verlies (of de vermindering van de lengte) van het Golgi-lint gepaard met verspreide, intacte, Golgi-ministapels. Deze situatie doet zich voor in een aantal celtypes en ook in modelsystemen, zoals de behandeling met nocodazole of de modulatie van de niveaus van TGN golgin GCC88. Membraantransport lijkt onder deze omstandigheden grotendeels onaangetast, met uitzondering van mogelijk grote lading (Ferraro et al., 2014; Lavieu et al., 2014), terwijl mTOR-signalering wordt verminderd en autofagie wordt versterkt (Gosavi et al., 2018).
(2) Verlies van zowel Golgi-lint als integriteit van Golgi-stapels. In dit scenario gaat de lintstructuur verloren en bovendien zijn cisternae van individuele stapels verminderd in lengte en/of aantal en kunnen ze ook geassocieerd zijn met gezwollen compartimenten, bijvoorbeeld de knock-out van GM130 (Liu et al., 2017) en mutaties van COG-subeenheden (Blackburn en Lupashin, 2016). Membraantransport en glycosylering worden waarschijnlijk beïnvloed in dit scenario, samen met verschillende signaleringsnetwerken.
(3) Verspreiding van één Golgi-compartiment. In dit scenario wordt slechts één Golgi-compartiment verstrooid, zoals de verstrooiing van de TGN onlangs gerapporteerde activering van het NLRP3 inflammasoom (Chen en Chen, 2018). De rest van de stapel/ribbon blijft onaangetast. Het identificeren van deze Golgi-structuren vereist het opnemen van EM- en optische microscopie en een aantal Golgi-markers over de stapel om de precieze veranderingen in Golgi-morfologie te definiëren.
(4) Conversie van Golgi-lint naar tubulovesiculaire elementen. In dit scenario worden zowel het Golgi-lint als de stapels uitgebreid verstoord met een dramatische toename van tubulovesiculaire structuren, zoals in het geval tijdens mitose (Wei en Seemann, 2017), verschillende geneesmiddelen en behandeling van cellen met amyloïde β (Joshi et al., 2014). Hier zou men een impact verwachten op veel van de functies van de Golgi.
Wij concluderen niet dat figuur 2 de enige morfologieën van Golgi “fragmenten” vertegenwoordigt en het is mogelijk dat aanvullende scenario’s zullen worden geïdentificeerd naarmate de structuren van Golgi “fragmenten” uitgebreider worden onderzocht.
Voorbeelden van een relatie tussen Golgi morfologie en celproces
De coördinatie van veranderingen in Golgi morfologie en verschillende celprocessen heeft veel aandacht gekregen. Voor een meer gedetailleerd overzicht van de achtergrondinformatie van de processen die beïnvloed worden door de veranderingen in Golgi-morfologie, zoals trafficking, glycosylatie, stress, DNA-reparatie, wordt de lezer verwezen naar een aantal recente reviews (Farhan en Rabouille, 2011; Millarte en Farhan, 2012; Sasaki en Yoshida, 2015; Gosavi en Gleeson, 2017; Makhoul et al., 2019). Hieronder volgen enkele voorbeelden om het bereik van celprocessen te benadrukken die worden gereguleerd of gecoördineerd door verschillende morfologische toestanden van de Golgi. In een aantal andere gevallen, zoals sommige kankers en stressresponsen, is het Golgi-lint verspreid als fragmenten, echter, de morfologie van deze Golgi-fragmenten is niet goed gekarakteriseerd. De bespreking hierin zal worden gericht op de voorbeelden waar de Golgi-morfologie goed is gedefinieerd.
DNA Repair and Cancer
Er is een intieme associatie tussen Golgi-morfologie en de DNA-schaderespons (Farber-Katz et al., 2014). De Golgi-membraanbinder, GOLPH3, is een oncogen en overexpressie van GOLPH3 resulteert in verbeterde celoverleving na DNA-schade (Scott et al., 2009; Farber-Katz et al., 2014). Omgekeerd, verlies van GOLPH3 voorkomt de dispersie van het Golgi-lint, versterkt het Golgi-lint en bevordert apoptose na DNA-schade. De DNA-schaderespons wordt gemedieerd door het kinase DNA-PK, dat GOLPH3 fosforyleert en Golgi-“fragmentatie” bevordert door actinepolymerisatie bij Golgi-membranen te versterken (Dippold et al., 2009). mTOR wordt gemoduleerd door de veranderingen in Golgi-morfologie gemedieerd door GOLPH3 (Scott et al., 2009) en draagt waarschijnlijk bij aan de uitkomst van de DNA-reactie. De precieze Golgi-structuur is dus verbonden met celoverleving en apoptose. De identiteit van de Golgi-structuren na de dispersie van het Golgi-lint door fosfo-GOLPH3 moet nog gekarakteriseerd worden. Toekomstige studies die de relatie tussen Golgi fragmentatie en mTOR signalisatie in kankercellen onderzoeken, zullen duidelijk de moeite waard zijn.
Genetische Aandoeningen Geassocieerd Met Veranderde Golgi Morfologie
Vele ziekten zijn geïdentificeerd met monogene aandoeningen veroorzaakt door overgeërfde mutaties van ofwel componenten geassocieerd met transport machines of van de enzymen die in de Golgi verblijven. Veel van deze ziekten zijn geassocieerd met fragmentatie van het Golgi-lint, bijvoorbeeld defecten in het geconserveerde oligomere Golgi-complex (COG) in aangeboren aandoeningen van de glycosylering (Miller en Ungar, 2012). In veel gevallen zijn de aandoeningen geassocieerd met pathologieën die beperkt zijn tot een beperkt aantal organen of weefsels. De basis voor weefselspecificiteit is slecht begrepen, maar waarschijnlijk te wijten aan tekortkomingen in glycosylatie en secretie en ook veranderingen in signaleringsnetwerken geassocieerd met het verlies van het Golgi-lint, zoals Golgi-stressreacties. De Golgi in alle COG subeenheid knock-out cellijnen vertonen matige tot ernstige veranderingen in morfologie zoals gekarakteriseerd door EM, geassocieerd met verlies van het lint, verwijde cisternae en in sommige gevallen verstoring van de mini-stacks. De toepassing van EM was fundamenteel bij het definiëren van de morfologische veranderingen (Blackburn en Lupashin, 2016).
Neurodegeneratieve Ziekten
De status van de Golgi in neurodegeneratieve ziekten heeft onlangs veel aandacht gekregen (figuur 1B). Verlies van het Golgi-lint is een gemeenschappelijk kenmerk van veel neurodegeneratieve ziekten, waaronder de ziekte van Alzheimer, de ziekte van Huntington, amyotrofische laterale sclerose en de ziekte van Parkinson (Gonatas et al., 2006; Haase en Rabouille, 2015; Rabouille en Haase, 2015; Sundaramoorthy et al., 2015). Het is zeer waarschijnlijk dat de verstoringen in de Golgi-architectuur bij deze ziekten bijdragen aan de pathologische processen. In de meeste gevallen is de precieze morfologische structuur van de Golgifragmenten in deze neurodegeneratieve ziekten niet gedefinieerd. Echter, twee experimentele systemen hebben onlangs onderzocht veranderingen in Golgi structuur en neuronale degeneratie in een aantal details. Ten eerste werd voorwaardelijke knock-out van GM130, een structurele golgin die het Golgi-lint reguleert, in het centrale zenuwstelsel aangetoond dat Golgi fragmentatie, atrofie van dendrieten en neuronale degeneratie in muizen veroorzaakt (Liu et al., 2017). EM analyse van de GM130 KO Purkinje cellen toonde een vermindering van de Golgi cisternal lengte en stapeling en bovendien waren de typische Golgi dendriet voorposten afwezig in deze GM130 -/- Purkinje cellen (Liu et al., 2017). In een tweede studie, hippocampale neuronen van muizen transgeen voor de Zweedse mutatie van amyloïde precursor eiwit (APP) en een mutant preseniline 1 subeenheid van γ-secretase, waar beide mutaties worden geassocieerd met de ziekte van Alzheimer op jonge leeftijd, onthulde uitgebreide Golgi fragmentatie door optische microscopie (Joshi et al., 2014). Kwantitatieve EM van neuronen in deze muizen toonde een vermindering van het aantal en de lengte van de cisternae in de stapels in vergelijking met neuronen van wild-type muizen. Bovendien waren de cisternae gezwollen. De veranderingen in Golgi morfologie in deze primaire neuronen werd aangetoond direct geassocieerd te zijn met het verhoogde niveau van amyloid-β productie. Primaire neuronen behandeld met synthetisch amyloid-β toonden ook een vergelijkbare fragmentatie van de Golgi en een toename van tubulovesiculaire structuren geassocieerd met Golgi cisternae in vergelijking met onbehandelde cellen. Het verlies van het Golgi-lint was te wijten aan de fosforylering van GRASP65, een Golgi-structuur-eiwit dat een sleutelrol speelt in de ontmanteling van het Golgi-lint en de stapels in mitose (Joshi et al., 2014). Vergelijking van deze twee studies hierboven is informatief, omdat de routes die de veranderingen in Golgi-morfologie bemiddelen in elk geval verschillen, wat leidt tot verschillen in de morfologie van de Golgi-“fragmenten”. Overwogen moet worden hoe deze verschillende paden die Golgi morfologie beïnvloeden de downstream reacties beïnvloeden.
mTOR Signaling
Ons lab heeft een experimentele benadering vastgesteld om het evenwicht tussen het Golgi-lint en Golgi mini-stapels te verstoren door het moduleren van de dosis van GCC88, een golgin gelegen aan de TGN. Deze strategie maakte het mogelijk een stabiele cellijn, HeLa-B6, tot stand te brengen die geen Golgi lint heeft. We hebben aangetoond dat GCC88 het evenwicht tussen Golgi-linten en mini-stapels reguleert door een actine-afhankelijk proces (Makhoul et al., 2019) en intersectin-1 (ITSN1) geïdentificeerd, een guanine nucleotide-uitwisselingsfactor voor cdc42 (Hussain et al., 2001), als een interactor van GCC88 die verantwoordelijk is voor het verlies van het Golgi-lint (Makhoul et al., 2019). Analyses van HeLa B6-cellen, die een Golgi-lint missen, toonden een verminderde mTOR-activiteit aan en een bijbehorende toename van autofagosomenbiogenese (Gosavi et al., 2018). mTOR is een van de belangrijkste signaalwegen van eukaryote cellen en staat bekend als een negatieve regulator van autofagie (Wullschleger et al., 2006). Vandaar dat het evenwicht van Golgi stapels naar Golgi lint een direct effect heeft op de mTORC1 pathway. Het gebruik van cis- en trans-compartiment specifieke markers, en EM tomografie, was van cruciaal belang bij het onthullen van de morfologische veranderingen in de Golgi ribbon.
Inflammation
Inflammasomes van het aangeboren immuunsysteem fungeren als een steiger voor caspase 1-afhankelijke activering van pro-inflammatoire cytokines (Broz en Dixit, 2016). Het NLRP3 (nucleotide-bindend domein, leucine-rijke-bevattende familie, pyrine-domein-bevattende-3) is een veelzijdig inflammasoom dat geactiveerd kan worden door een reeks microbiële en niet-microbiële stimuli wat resulteert in secretie van pro-inflammatoire cytokines interleukine 1β (IL-1β) en interleukine 18 (IL-18) en geprogrammeerde celdood door pyroptose. Een recente studie heeft aangetoond dat de activering van het cytosolische NLRP3 door stimuli gepaard gaat met rekrutering van NLRP3 naar verspreide TGN membranen om NLRP3 scaffold assemblage te vergemakkelijken (Chen en Chen, 2018). De verspreide TGN, maar niet de onderliggende andere compartimenten, rekruteert specifiek NLRP3, via PI4P, om het downstream adaptorcomplex ASC te assembleren dat polymerisatie ondergaat in de peri-nucleaire regio voordat caspase-1 wordt gerekruteerd om de downstream signaalroute te activeren (Chen en Chen, 2018). Deze belangrijke bevinding toont dus aan dat de modulatie van de architectuur van de TGN selectief, cruciaal is in de activering van deze signaalweg. Het gebruik van compartimentspecifieke markers (TGN en cis-Golgi) samen met optische en EM-analyse was van cruciaal belang bij het onthullen van de morfologische veranderingen in het Golgi-lint.
Conclusieve opmerkingen
De precieze architectuur van de Golgi-morfologie wordt bepaald door de hoge resolutie optische microscopie en door EM. Het opnemen van deze informatie in toekomstige studies in het veld zal een aanzienlijk inzicht geven in de dynamiek van de Golgi, de routes voor verstoring van de lintstructuur en de functionele gevolgen geassocieerd met deze verschillende routes.
Samenvattend stellen wij voor dat het gebruik van de term “gefragmenteerde Golgi” ontoereikend is om Golgi-structuren geassocieerd met vele behandelingen en aandoeningen te beschrijven en dat de verschillen in deze Golgi-structuren waarschijnlijk fysiologisch relevant zullen zijn. Gezien de dynamische aard van het Golgi-apparaat, is het mogelijk dat er een evenwicht is tussen verschillende morfologische toestanden van de Golgi op een gegeven moment, dat wil zeggen, mini-stapels en lintstructuren. Inzicht in het dynamische evenwicht tussen de verschillende Golgi-morfologieën in moleculair detail is van cruciaal belang voor een volledige beoordeling van dit organel tijdens normale celprocessen en ook onder pathologische omstandigheden. Het zal fascinerend zijn om te zien wat zich ontvouwt als we meer leren over de cel sensing functies van deze complexe organel en de relatie tussen deze functies en Golgi structuren.
Data Beschikbaarheid
Alle datasets geanalyseerd voor deze studie zijn opgenomen in het manuscript en de aanvullende bestanden.
Author Contributions
Alle vermelde auteurs hebben een substantiële, directe en intellectuele bijdrage geleverd aan het werk, en goedgekeurd voor publicatie.
Funding
Dit werk werd ondersteund door financiering van de Australian Research Council (DP160102394).
Conflict of Interest Statement
De auteurs verklaren dat het onderzoek werd uitgevoerd in de afwezigheid van enige commerciële of financiële relaties die zouden kunnen worden opgevat als een potentieel belangenconflict.
Afkortingen
GM130, cis-Golgi matrix proteïne; GMAP210, Golgi microtubule associated protein; GOLPH3, Golgi phosphoprotein 3; GRASP, Golgi reassembly stacking protein; MTOC, microtubule organizing center; mTOR, mechanistic target of rapamycin; PI4P, phosphatidylinositol-4-phosphate; TGN, trans-Golgi network.
Abdel Rahman, A. M., Ryczko, M., Nakano, M., Pawling, J., Rodrigues, T., Johswich, A., et al. (2015). Golgi N-glycan vertakkingen N-acetylglucosaminyltransferases I, V en VI bevorderen nutriëntenopname en metabolisme. Glycobiology 25, 225-240. doi: 10.1093/glycob/cwu105
PubMed Abstract | CrossRef Full Text | Google Scholar
Blackburn, J. B., and Lupashin, V. V. (2016). Het creëren van knock-outs van geconserveerde oligomere golgi-complex subeenheden met behulp van CRISPR-gemedieerde gen editing gekoppeld aan een selectie strategie op basis van glycosylatie defecten geassocieerd met een verminderde COG-complex functie. Methods Mol. Biol. 1496, 145-161. doi: 10.1007/978-1-4939-6463-5_12
PubMed Abstract | CrossRef Full Text | Google Scholar
Boncompain, G., and Perez, F. (2013). De vele routes van Golgi-afhankelijke trafficking. Histochem. Cell Biol. 140, 251-260. doi: 10.1007/s00418-013-1124-7
PubMed Abstract | CrossRef Full Text | Google Scholar
Broz, P., and Dixit, V. M. (2016). Inflammasomes: mechanisme van assemblage, regulatie en signalering. Nat. Rev. Immunol. 16, 407-420. doi: 10.1038/nri.2016.58
PubMed Abstract | CrossRef Full Text | Google Scholar
Chen, J., and Chen, Z. J. (2018). PtdIns4P op verspreide trans-Golgi netwerk medieert NLRP3 inflammasoom activatie. Nature 564, 71-76. doi: 10.1038/s41586-018-0761-3
PubMed Abstract | CrossRef Full Text | Google Scholar
Chia, J., Goh, G., Racine, V., Ng, S., Kumar, P., and Bard, F. (2012). RNAi screening onthult een groot signaleringsnetwerk dat het Golgi-apparaat in menselijke cellen controleert. Mol. Syst. Biol. 8:629. doi: 10.1038/msb.2012.59
PubMed Abstract | CrossRef Full Text | Google Scholar
Corda, D., Barretta, M. L., Cervigni, R. I., and Colanzi, A. (2012). Golgi-complex fragmentatie in G2/M overgang: een organel-gebaseerd celcyclus checkpoint. IUBMB Life 64, 661-670. doi: 10.1002/iub.1054
PubMed Abstract | CrossRef Full Text | Google Scholar
De Matteis, M. A., Mironov, A. A., and Beznoussenko, G. V. (2008). De Golgi lint en de functie van de Golgins. New York, NY: Springer-Verlag/Wein.
Google Scholar
Dippold, H. C., Ng, M. M., Farber-Katz, S. E., Lee, S. K., Kerr, M. L., Peterman, M. C., et al. (2009). GOLPH3 slaat een brug tussen fosfatidylinositol-4-fosfaat en actomyosine om de Golgi op te rekken en vorm te geven en zo de knopvorming te bevorderen. Cell 139, 337-351. doi: 10.1016/j.cell.2009.07.052
PubMed Abstract | CrossRef Full Text | Google Scholar
Efimov, A., Kharitonov, A., Efimova, N., Loncarek, J., Miller, P. M., Andreyeva, N., et al. (2007). Asymmetrische CLASP-afhankelijke nucleatie van niet-centrosomale microtubuli in het trans-Golgi netwerk. Dev. Cell 12, 917-930. doi: 10.1016/j.devcel.2007.04.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Farber-Katz, S. E., Dippold, H. C., Buschman, M. D., Peterman, M. C., Xing, M., Noakes, C. J., et al. (2014). DNA schade triggert Golgi dispersie via DNA-PK en GOLPH3. Cell 156, 413-427. doi: 10.1016/j.cell.2013.12.023
PubMed Abstract | CrossRef Full Text | Google Scholar
Farhan, H., and Rabouille, C. (2011). Signalen naar en van de secretorische pathway. J. Cell Sci. 124, 171-180. doi: 10.1242/jcs.076455
PubMed Abstract | CrossRef Full Text | Google Scholar
Farhan, H., Wendeler, M. W., Mitrovic, S., Fava, E., Silberberg, Y., Sharan, R., et al. (2010). MAPK signaling to the early secretory pathway revealed by kinase/phosphatase functional screening. J. Cell Biol. 189, 997-1011. doi: 10.1083/jcb.200912082
PubMed Abstract | CrossRef Full Text | Google Scholar
Ferraro, F., Kriston-Vizi, J., Metcalf, D. J., Martin-Martin, B., Freeman, J., Burden, J. J., et al. (2014). Een tweelagige Golgi-gebaseerde controle van organelgrootte onderbouwt de functionele plasticiteit van endotheelcellen. Dev. Cell 29, 292-304. doi: 10.1016/j.devcel.2014.03.021
PubMed Abstract | CrossRef Full Text | Google Scholar
Gonatas, N. K., Stieber, A., and Gonatas, J. O. (2006). Fragmentatie van het Golgi-apparaat in neurodegeneratieve ziekten en celdood. J. Neurol. Sci. 246, 21-30. doi: 10.1016/j.jns.2006.01.019
PubMed Abstract | CrossRef Full Text | Google Scholar
Gosavi, P., and Gleeson, P. A. (2017). De functie van de Golgi lintstructuur- Een blijvende mytery ontvouwt zich! Bioessays 39:1700063. doi: 10.1002/bies.201700063
PubMed Abstract | CrossRef Full Text | Google Scholar
Gosavi, P., Houghton, F. J., Mcmillan, P. J., Hanssen, E., and Gleeson, P. A. (2018). Het Golgi-lint in zoogdiercellen reguleert autofagie negatief door de mTOR-activiteit te moduleren. J. Cell Sci. 131:jcs211987. doi: 10.1242/jcs.211987
PubMed Abstract | CrossRef Full Text | Google Scholar
Gunn, P. A., Gliddon, B. L., Londrigan, S. L., Lew, A. M., Van Driel, I. R., and Gleeson, P. A. (2011). Het Golgi-apparaat in de endomembraanrijke gastrische pariëtale cellen bestaat als functionele stabiele mini-stapels verspreid over het cytoplasma. Biol. Cell 103, 559-572. doi: 10.1042/BC20110074
PubMed Abstract | CrossRef Full Text | Google Scholar
Haase, G., and Rabouille, C. (2015). Golgifragmentatie in ALS-motorneuronen. Nieuwe mechanismen gericht op microtubuli, tethers, en transport vesicles. Front. Neurosci. 9:448. doi: 10.3389/fnins.2015.00448
PubMed Abstract | CrossRef Full Text | Google Scholar
Hussain, N. K., Jenna, S., Glogauer, M., Quinn, C. C., Wasiak, S., Guipponi, M., et al. (2001). Endocytisch eiwit intersectin-l reguleert actine-assemblage via Cdc42 en N-WASP. Nat. Cell Biol. 3, 927-932. doi: 10.1038/ncb1001-927
PubMed Abstract | CrossRef Full Text | Google Scholar
Joshi, G., Chi, Y., Huang, Z., and Wang, Y. (2014). Abeta-geïnduceerde Golgi-fragmentatie bij de ziekte van Alzheimer verhoogt de Abeta productie. Proc. Natl. Acad. Sci. U.S.A. 111, E1230-E1239. doi: 10.1073/pnas.1320192111
PubMed Abstract | CrossRef Full Text | Google Scholar
Kreft, M. E., Di Giandomenico, D., Beznoussenko, G. V., Resnik, N., Mironov, A. A., and Jezernik, K. (2010). Golgi-apparaat fragmentatie als een mechanisme verantwoordelijk voor uniforme levering van uroplakines aan de apicale plasmamembraan van uroepitheliale cellen. Biol. Cell 102, 593-607. doi: 10.1042/bc20100024
PubMed Abstract | CrossRef Full Text | Google Scholar
Kupfer, A., Dennert, G., and Singer, S. J. (1983). Polarization of the Golgi apparatus and the microtubule-organizing center within cloned natural killer cells bound to their targets. Proc. Natl. Acad. Sci. U.S.A. 80, 7224-7228. doi: 10.1073/pnas.80.23.7224
PubMed Abstract | CrossRef Full Text | Google Scholar
Lamb, C. A., Yoshimori, T., and Tooze, S. A. (2013). De autofagosoom: oorsprong onbekend, biogenese complex. Nat. Rev. Mol. Cell Biol. 14, 759-774. doi: 10.1038/nrm3696
PubMed Abstract | CrossRef Full Text | Google Scholar
Lasiecka, Z. M., and Winckler, B. (2011). Mechanismen van gepolariseerde membraantransport in neuronen – focus op endosomen. Mol. Cell. Neurosci. 48, 278-287. doi: 10.1016/j.mcn.2011.06.013
PubMed Abstract | CrossRef Full Text | Google Scholar
Lavieu, G., Dunlop, M. H., Lerich, A., Zheng, H., Bottanelli, F., and Rothman, J. E. (2014). De Golgi lintstructuur vergemakkelijkt anterograde transport van grote ladingen. Mol. Biol. Cell 25, 3028-3036. doi: 10.1091/mbc.E14-04-093
PubMed Abstract | CrossRef Full Text | Google Scholar
Lazaro-Dieguez, F., Jimenez, N., Barth, H., Koster, A. J., Renau-Piqueras, J., Llopis, J. L., et al. (2006). Actin filaments are involved in the maintenance of Golgi cisternae morphology and intra-Golgi pH. Cell Motil. Cytoskelet 63, 778-791. doi: 10.1002/cm.20161
PubMed Abstract | CrossRef Full Text | Google Scholar
Liu, C., Mei, M., Li, Q., Roboti, P., Pang, Q., Ying, Z., et al. (2017). Verlies van de golgin GM130 veroorzaakt Golgi verstoring, Purkinje neuron verlies, en ataxie in muizen. Proc. Natl. Acad. Sci. U.S.A. 114, 346-351. doi: 10.1073/pnas.1608576114
PubMed Abstract | CrossRef Full Text | Google Scholar
Lu, Z., Joseph, D., Bugnard, E., Zaal, K. J., and Ralston, E. (2001). Golgi complex reorganisatie tijdens spier differentiatie: visualisatie in levende cellen en mechanisme. Mol. Biol. Cell 12, 795-808. doi: 10.1091/mbc.12.4.795
PubMed Abstract | CrossRef Full Text | Google Scholar
Luini, A., and Parashuraman, S. (2016). Signaling bij de Golgi: sensing and controlling the membrane fluxes. Curr. Opin. Cell Biol. 39, 37-42. doi: 10.1016/j.ceb.2016.01.014
PubMed Abstract | CrossRef Full Text | Google Scholar
Makhoul, C., Gosavi, P., Duffield, R., Delbridge, B., Williamson, N. A., and Gleeson, P. A. (2019). Intersectin-1 interacteert met de golgin GCC88 om het actinenetwerk en de Golgi-architectuur te koppelen. Mol. Biol. Cell 30, 370-386. doi: 10.1091/mbc.E18-05-0313
PubMed Abstract | CrossRef Full Text | Google Scholar
Makhoul, C., Gosavi, P., and Gleeson, P. A. (2018). De Golgi architectuur en cel sensing. Biochem. Soc. Trans. 46, 1063-1072. doi: 10.1042/BST20180323
PubMed Abstract | CrossRef Full Text | Google Scholar
Mayinger, P. (2011). Signalering bij de Golgi. Cold Spring Harb. Perspect. Biol. 3:a005314.
Google Scholar
Millarte, V., Boncompain, G., Tillmann, K., Perez, F., Sztul, E., and Farhan, H. (2015). Fosfolipase C gamma1 reguleert vroege secretoire trafficking en celmigratie via interactie met p115. Mol. Biol. Cell 26, 2263-2278. doi: 10.1091/mbc.E15-03-0178
PubMed Abstract | CrossRef Full Text | Google Scholar
Millarte, V., and Farhan, H. (2012). De Golgi in celmigratie: regulatie door signaaltransductie en de implicaties voor kankercelmetastase. ScientificWorldJournal 2012:498278. doi: 10.1100/2012/498278
PubMed Abstract | CrossRef Full Text | Google Scholar
Miller, V. J., and Ungar, D. (2012). Re’COG’nition at the Golgi. Traffic 13, 891-897. doi: 10.1111/j.1600-0854.2012.01338.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Petrosyan, A. (2015). Onco-Golgi: is fragmentatie een poort naar kankerprogressie? Biochem. Mol. Biol. J. 1:16.
PubMed Abstract | Google Scholar
Puthenveedu, M. A., Bachert, C., Puri, S., Lanni, F., and Linstedt, A. D. (2006). GM130 en GRASP65-afhankelijke laterale cisternale fusie maakt uniforme Golgi-enzym distributie mogelijk. Nat. Cell Biol. 8, 238-248. doi: 10.1038/ncb1366
PubMed Abstract | CrossRef Full Text | Google Scholar
Rabouille, C., and Haase, G. (2015). Editorial: golgi pathologie in neurodegeneratieve ziekten. Front. Neurosci. 9:489. doi: 10.3389/fnins.2015.00489
PubMed Abstract | CrossRef Full Text | Google Scholar
Rabouille, C., and Kondylis, V. (2007). Golgi ribbon unlinking: een organel-gebaseerd G2/M checkpoint. Cell Cycle 6, 2723-2729. doi: 10.4161/cc.6.22.4896
PubMed Abstract | CrossRef Full Text | Google Scholar
Rambourg, A., and Clermont, Y. (1997). Three-Dimensional Structure of the Golgi Apparatus in Mammalian Cells. Basel: Birkhauser.
Google Scholar
Sanders, A. A., and Kaverina, I. (2015). Nucleatie en dynamiek van Golgi-afgeleide microtubuli. Front. Neurosci. 9:431. doi: 10.3389/fnins.2015.00431
CrossRef Full Text | Google Scholar
Sasaki, K., and Yoshida, H. (2015). Organelle autoregulation-stress responses in the ER, Golgi, mitochondria and lysosome. J. Biochem. 157, 185-195. doi: 10.1093/jb/mvv010
PubMed Abstract | CrossRef Full Text | Google Scholar
Scott, K. L., Kabbarah, O., Liang, M. C., Ivanova, E., Anagnostou, V., Wu, J., et al. (2009). GOLPH3 moduleert mTOR signalering en rapamycine gevoeligheid in kanker. Nature 459, 1085-1090. doi: 10.1038/nature08109
PubMed Abstract | CrossRef Full Text | Google Scholar
Serebrenik, Y. V., Hellerschmied, D., Toure, M., Lopez-Giraldez, F., Brookner, D., and Crews, C. M. (2018). Gerichte ontvouwing van eiwitten legt een Golgi-specifieke transcriptionele stressrespons bloot. Mol. Biol. Cell 29, 1284-1298. doi: 10.1091/mbc.E17-11-0693
PubMed Abstract | CrossRef Full Text | Google Scholar
Sundaramoorthy, V., Sultana, J. M., and Atkin, J. D. (2015). Golgi fragmentatie in amyotrofische laterale sclerose, een overzicht van mogelijke triggers en gevolgen. Front. Neurosci. 9:400. doi: 10.3389/fnins.2015.00400
PubMed Abstract | CrossRef Full Text | Google Scholar
Wei, J. H., and Seemann, J. (2010). Het ontrafelen van de Golgi lint. Traffic 11, 1391-1400. doi: 10.1111/j.1600-0854.2010.01114.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Wei, J. H., and Seemann, J. (2017). Golgi ribbon disassembly during mitosis, differentiation and disease progression. Curr. Opin. Cell Biol. 47, 43-51. doi: 10.1016/j.ceb.2017.03.008
PubMed Abstract | CrossRef Full Text | Google Scholar
Wu, J., De Heus, C., Liu, Q., Bouchet, B. P., Noordstra, I., Jiang, K., et al. (2016). Molecular pathway of microtubule organization at the Golgi apparatus. Dev. Cell 39, 44-60. doi: 10.1016/j.devcel.2016.08.009
PubMed Abstract | CrossRef Full Text | Google Scholar
Wullschleger, S., Loewith, R., and Hall, M. N. (2006). TOR signalering in groei en metabolisme. Cell 124, 471-484.
Google Scholar
Yamamoto, H., Kakuta, S., Watanabe, T. M., Kitamura, A., Sekito, T., Kondo-Kakuta, C., et al. (2012). Atg9 vesicles are an important membrane source during early steps of autophagosome formation. J. Cell Biol. 198, 219-233. doi: 10.1083/jcb.201202061
PubMed Abstract | CrossRef Full Text | Google Scholar
Zappa, F., Failli, M., and De Matteis, M. A. (2018). Het Golgi-complex in ziekte en therapie. Curr. Opin. Cell Biol. 50, 102-116. doi: 10.1016/j.ceb.2018.03.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Leave a Reply