Frontiers in Celland Developmental Biology
Baggrund
Golgi-apparatets grundlæggende enhed anses normalt for at være en stak af cisterner, som er stærkt polariseret, med cis-siden, der modtager last fra ER, og trans-siden af stakken, TGN, der er forbundet med sortering af last til post-Golgi-eksport (Boncompain og Perez, 2013). Organiseringen af disse mini-stakke i cellen varierer imidlertid blandt forskellige organismer. I planter og hvirvelløse dyr er de enkelte Golgi-stakke spredt uafhængigt af hinanden i hele cytoplasmaet, mens de enkelte Golgi-stakke i de fleste hvirveldyrceller under interfasen er smeltet sammen til en kompakt båndstruktur, der er placeret tæt på MTOC (Wei og Seemann, 2017). Strukturen af Golgi-båndet i pattedyrceller afsløres bedst ved elektronmikroskopi (Rambourg og Clermont, 1997); optisk mikroskopi med høj opløsning ved hjælp af cis- og trans-markører kan også detektere båndorganisationen (Gosavi et al., 2018). Et vigtigt spørgsmål, der undersøges i feltet, er relevansen af Golgi-båndsstrukturen og de funktionelle forskelle i Golgi-båndet sammenlignet med “isolerede Golgi-ministakke” eller andre tilstande af Golgi-arkitektur. Med andre ord, hvad er den evolutionære fordel ved den mere komplekse båndmorfologi af Golgi-apparatet i hvirveldyrceller, og hvilke funktioner kan reguleres ved en overgang til en ikke-båndstruktur?
Studier i de seneste par år afslører, at Golgi-membraner udgør en platform for regulering af en række cellulære processer, herunder cellepolarisering (Kupfer et al, 1983), rettet migration (Millarte og Farhan, 2012), stress (Sasaki og Yoshida, 2015), DNA-reparation (Farber-Katz et al., 2014), mitose (Rabouille og Kondylis, 2007), metabolisme (Abdel Rahman et al., 2015), pro-inflammatoriske reaktioner (Chen og Chen, 2018) og autofagi (Yamamoto et al., 2012; Lamb et al., 2013). Faktisk er der i pattedyrceller nu betydelige beviser for, at Golgi, ligesom andre intracellulære organeller, kan fungere som en cellesensor (Farhan og Rabouille, 2011; Mayinger, 2011; Millarte og Farhan, 2012; Sasaki og Yoshida, 2015; Luini og Parashuraman, 2016; Gosavi og Gleeson, 2017; Makhoul et al., 2018). Det er også ved at blive klart, at den præcise morfologi af Golgi er relevant for reguleringen af en række af disse celleprocesser (Makhoul et al., 2018). Associeringen af Golgi-morfologi med signalering blev båret af en genom bred kinom- og fosfatom-screening, som identificerede en stor kohorte af kinaser og fosfataser (20% af det samlede antal i genomet), der påvirkede Golgis morfologi (Chia et al., 2012). Ændringerne i Golgi-morfologien omfattede enten fragmentering af Golgi (tab af Golgi-båndet) eller dannelse af en meget kompakt og kondenseret Golgi i den perinukleære placering. Aktinets relevans i reguleringen af Golgi-morfologien blev fremhævet i denne undersøgelse ved identifikation af en række kinaser, f.eks. ROCK1 og PAK1, som regulerer aktindynamikken og modulerer Golgi-strukturen (Chia et al., 2012). Andre genom brede analyser har også fremhævet sandsynligheden for, at Golgi kan modtage og transmittere en lang række signaler, der kan påvirke ikke kun membrantransportveje, men også andre processer, apoptose, mitose, autofagi og stressresponser (Farhan et al., 2010; Millarte et al., 2015).
Golgi-båndstrukturen er meget dynamisk og kan undergå meget hurtig remodellering under en række forskellige forhold. For eksempel under mitose er adskillelsen af Golgi-båndet en tidlig begivenhed i G2/M-overgangen og spiller en vigtig rolle som et kontrolpunkt i cellecyklus ved at fremme mitotisk indgang (Wei og Seemann, 2010; Corda et al., 2012). Reguleringen af Golgi-dynamikken er medieret af interaktioner mellem molekylære stilladser placeret på Golgimembranen og cytoskelettet. MT-dynamikken kan regulere placeringen af Golgi-båndet ved centrosomet og omplaceringen af Golgi for at lette polariseret trafikering og målrettet sekretion (Millarte og Farhan, 2012; Sanders og Kaverina, 2015). Derudover kan membrankomponenter i Golgi også kerneopbygge og stabilisere mikrotubuli ved både cis- og trans-Golgi, og derfor er Golgi selv også en MTOC (Efimov et al., 2007; Wu et al., 2016). Actin-medierede processer bidrager også til Golgis form og funktion, og mindst ni Golgi-lokaliserede molekylære stilladser er blevet identificeret, som interagerer med actin-cytoskelettet . Forøgelse af actinpolymerisation ved Golgi resulterer i spredning af båndet, mens hæmning af actinpolymerisation med specifikke lægemidler såsom latrunculin A resulterer i komprimering af Golgi (Lazaro-Dieguez et al., 2006; Makhoul et al., 2018, 2019). I betragtning af disse reguleringsnetværk, der modulerer Golgi-båndets struktur, anser vi det for meget plausibelt, at balancen mellem Golgi-båndet og Golgi-mini-stakkene kan definere både de kvalitative og kvantitative svar af signalveje. Forbindelsen mellem Golgi-morfologi og signalering har også vigtige forgreninger for forståelsen af det molekylære grundlag for en række sygdomme, der er forbundet med tab af Golgi-båndet og fremkomsten af et spredt Golgi. For eksempel har det vist sig, at overlevelsen af nogle kræftceller er forbundet med en spredt Golgi, som reducerer niveauet af apoptose (Farber-Katz et al., 2014; Petrosyan, 2015).
Hvad betyder Golgi-fragmentering?
Udtrykket “Golgi-fragmentering” bruges almindeligvis til at beskrive den morfologiske status af en spredt Golgi i pattedyrceller, som detekteret ved optisk mikroskopi farvet med Golgi-markører. En spredt Golgi observeres ofte i eksperimentelle systemer, for eksempel når celler behandles med stoffer, der forstyrrer cytoskelettet, f.eks. nocodazol (Wei og Seemann, 2010), når membranflux forstyrres, eller når komponenter af Golgi-transportmaskineriet eller cytoskeletets interaktionssystem slås ned, slås ud eller overeksprimeres (Zappa et al., 2018). I forskellige fysiologiske tilstande, for eksempel stress (Serebrenik et al., 2018) og patologiske tilstande, især kræft og neurodegeneration, har Golgi ofte mistet den typiske kompakte juxtanucleære placering og observeres ved konfokal mikroskopi som spredte strukturer i hele cytoplasmaet . Der er et stigende antal rapporter, der beskriver Golgi-fragmentering under fysiologiske og patologiske forhold (figur 1A). Et problem ved brugen af udtrykket “fragmenteret” er imidlertid, at det indebærer, at Golgi’s strukturelle integritet er tabt, og at den morfologi, der er forbundet med den “fragmenterede” struktur, repræsenterer en opløst, unormal eller ødelagt organel. I en række tilfælde er dette klart misvisende, da de enkelte Golgi-stakke kan forblive intakte og kan opretholde organellens klassiske funktioner, nemlig glykosylering og membrantransport. Der forekommer faktisk spredte Golgi-ministakke i nogle specialiserede celler, f.eks. differentierede myoblaster (Lu et al., 2001), differentierede neuroner, som indeholder individuelle Golgi-stakke eller “forposter” langs dendritter (Lasiecka og Winckler, 2011), gastriske parietalceller (Gunn et al., 2011) og uroepithelceller i urinblæren (Kreft et al., 2010), uden at der tilsyneladende er mangel på membrantransport og glykosylering. Spredningen af Golgi-båndet i forbindelse med eksperimentelle og patologiske forhold kan snarere afspejle en ændring af den dynamiske balance mellem den kompakte båndmorfologi og de individuelle Golgi-ministakke eller kan resultere i en forstyrrelse af båndarkitekturen såvel som Golgi-stakkenes integritet. Dette er et relevant spørgsmål, da Golgis præcise morfologiske status højst sandsynligt vil påvirke effektiviteten af transport og glykosylering i nogle tilfælde (Puthenveedu et al., 2006) og i andre tilfælde en række signalnetværk, men ikke nødvendigvis transport eller glykosylering. Det er vigtigt at skelne mellem intakte Golgi-ministakke og tab af Golgistakkernes integritet, når man skal definere, hvad der menes med en “fragmenteret” Golgi. Her gennemgår vi de forskellige Golgi-morfologier, der er blevet påvist og karakteriseret i eksperimentelle, fysiologiske og patologiske omgivelser.
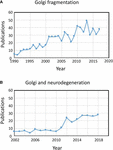
Figur 1. Publikationer, der identificerer fragmentering af Golgi-båndet. (A) Antal publikationer pr. år med udtrykket Golgi-fragmentering i enten titlen eller i resuméet. (B) Antal publikationer pr. år, hvor Golgi er blevet undersøgt i forbindelse med neurodegenerative sygdomme. Data er fra Alexandru Dan Corlan. Medline trend: automatiseret årlig statistik over PubMed-resultater for alle forespørgsler, 2004. Webressource på URL: http://dan.corlan.net/medline-trend.html. Tilgået: 2019-04-29.
Rethinking the Terminology of Golgi Morphological States
Strukturerne af Golgi-fragmenterne varierer afhængigt af arten af den vej, der er involveret for at forstyrre eller modulere Golgi-båndet. Det er vigtigt at få en bedre ultrastrukturel karakterisering af Golgi-“fragmenterne” efter tab af Golgi-båndet, da det funktionelle resultat sandsynligvis vil være meget forskelligt afhængigt af de præcise Golgi-strukturer. Ud over de forhold, der resulterer i et forlænget Golgi-bånd, kan vi ud fra litteraturen identificere mindst fire forskellige scenarier, der er forbundet med Golgi-båndets “fragmentering”. Disse er afbildet i figur 2 og beskrevet som følger:
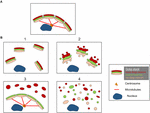
Figur 2. Model, der viser forskellige Golgi-morfologier efter “fragmentering” af Golgi-båndstrukturen. (A) Intakt Golgi-båndsstruktur og (B) forskellige scenarier, der viser tab af Golgi-båndet. (1) Et scenarie, hvor intakte Golgi-ministakke er spredt ud over hele cytoplasmaet; (2) Et scenarie, hvor integriteten af de spredte Golgi-stakke er kompromitteret med forkortede cisterner, hævelse af cisterner og forøgelse af Golgi-associerede tubuli og vesikler; (3) Et scenarie, hvor der er spredning af et Golgi-kompartment. Her spredes TGN selektivt i hele cytoplasmaet, mens resten af stakken forbliver i en båndstruktur. (4) Scenarie, hvor der er tab af bånd og stakke med Golgi-membraner spredt overvejende som tubuli og vesikler. Numrene henviser til klassifikationen af Golgi-morfologierne i teksten.
(1) Omdannelse af Golgi-bånd til Golgi-stakke. Her er tabet (eller reduktionen af længden) af Golgi-båndet forbundet med spredte, intakte Golgi-mini-stakke. Denne situation forekommer i en række celletyper og også i modelsystemer, f.eks. ved behandling med nocodazol eller ved modulation af niveauerne af TGN golgin GCC88. Membrantransport synes stort set ikke at være påvirket under disse forhold, med undtagelse af muligvis stor last (Ferraro et al., 2014; Lavieu et al., 2014), mens mTOR-signalering reduceres og autofagi øges (Gosavi et al., 2018).
(2) Tab af både Golgi-bånd og integritet af Golgi-stakke. I dette scenarie er båndstrukturen tabt, og desuden er cisternerne i individuelle stakke reduceret i længde og/eller antal og kan også være forbundet med opsvulmede rum, for eksempel knockout af GM130 (Liu et al., 2017) og mutationer af COG-underenheder (Blackburn og Lupashin, 2016). Membrantransport og glykosylering vil sandsynligvis blive påvirket i dette scenarie sammen med forskellige signalnetværk.
(3) Spredning af et Golgi-kompartment. I dette scenarie spredes kun ét Golgi-kompartment, som f.eks. spredningen af TGN, der for nylig blev rapporteret om aktivering af NLRP3-inflammasomet (Chen og Chen, 2018). Resten af stakken/ribbon forbliver upåvirket. Identifikation af disse Golgi-strukturer kræver inkorporering af EM og optisk mikroskopi og en række Golgi-markører på tværs af stakken for at definere de præcise ændringer i Golgi-morfologien.
(4) Omdannelse af Golgi-bånd til tubulovesikulære elementer. I dette scenarie forstyrres både Golgi-båndet og stakken i vid udstrækning med dramatiske stigninger i tubulovesikulære strukturer, som i tilfældet under mitose (Wei og Seemann, 2017), forskellige lægemidler og behandling af celler med amyloid β (Joshi et al., 2014). Her ville man forvente en indvirkning på mange af Golgis funktioner.
Vi udleder ikke, at Figur 2 repræsenterer de eneste morfologier af Golgi-“fragmenter”, og det er muligt, at yderligere scenarier vil blive identificeret, efterhånden som strukturerne af Golgi-“fragmenter” undersøges mere indgående.
Eksempler på en sammenhæng mellem Golgi-morfologi og celleproces
Koordineringen af ændringer i Golgi-morfologi og forskellige celleprocesser har fået betydelig opmærksomhed. For en mere detaljeret sammenfatning af baggrundsoplysningerne om de processer, der påvirkes af ændringerne i Golgi-morfologi, såsom trafikering, glykosylering, stress, DNA-reparation, henvises læseren til en række nyere oversigter (Farhan og Rabouille, 2011; Millarte og Farhan, 2012; Sasaki og Yoshida, 2015; Gosavi og Gleeson, 2017; Makhoul et al., 2019). Nedenfor er der nogle eksempler for at fremhæve den række af celleprocesser, der reguleres eller koordineres af forskellige morfologiske tilstande af Golgi. I en række andre tilfælde, såsom visse kræftformer og stressreaktioner, er Golgi-båndet spredt som fragmenter, men morfologien af disse Golgi-fragmenter er ikke blevet godt karakteriseret. Diskussionen heri vil være fokuseret på de eksempler, hvor Golgi-morfologien er veldefineret.
DNA-reparation og kræft
Der er en tæt sammenhæng mellem Golgi-morfologi og DNA-skadesrespons (Farber-Katz et al., 2014). Golgimembranens tether, GOLPH3, er en onkogen, og overekspression af GOLPH3 resulterer i øget celleoverlevelse efter DNA-skader (Scott et al., 2009; Farber-Katz et al., 2014). Omvendt forhindrer tab af GOLPH3 spredningen af Golgi-båndet, forbedrer Golgi-båndet og fremmer apoptose efter DNA-skader. DNA-skadesresponset medieres af kinasen DNA-PK, som fosforylerer GOLPH3 og fremmer Golgi-“fragmentering” ved at øge aktinpolymerisationen ved Golgimembranerne (Dippold et al., 2009). mTOR moduleres af de ændringer i Golgi-morfologien, der medieres af GOLPH3 (Scott et al., 2009) og bidrager sandsynligvis til resultatet af DNA-responset. Derfor er den præcise Golgi-struktur bundet til celleoverlevelse og apoptose. Identiteten af Golgi-strukturerne efter spredningen af Golgi-båndet ved hjælp af phospho-GOLPH3 mangler stadig at blive karakteriseret. Det er klart, at fremtidige undersøgelser, der undersøger forholdet mellem Golgi-fragmentering og mTOR-signalering i kræftceller, vil være meget værdifulde.
Genetiske lidelser forbundet med ændret Golgi-morfologi
Mange sygdomme er blevet identificeret med monogene lidelser forårsaget af arvelige mutationer af enten komponenter forbundet med transportmaskineriet eller af de enzymer, der bor i Golgi. Mange af disse sygdomme er forbundet med fragmentering af Golgi-båndet, f.eks. defekter i det konserverede oligomeriske Golgi-kompleks (COG) i medfødte glykosyleringsforstyrrelser (Miller og Ungar, 2012). I mange tilfælde er lidelserne forbundet med patologier, der er begrænset til et begrænset antal organer eller væv. Grundlaget for vævsspecificitet er dårligt forstået, men skyldes sandsynligvis mangler i glykosylering og sekretion og også ændringer i signalnetværk, der er forbundet med tabet af Golgi-båndet, såsom Golgi-stressresponser. Golgi i alle cellelinjer med knockout af COG-underenheden viser moderate til alvorlige morfologiske ændringer, som er karakteriseret ved EM, og som er forbundet med tab af båndet, udvidede cisterner og i nogle tilfælde afbrydelse af mini-stacks. Anvendelsen af EM var grundlæggende for at definere de morfologiske ændringer (Blackburn og Lupashin, 2016).
Neurodegenerative sygdomme
Status af Golgi i neurodegenerative sygdomme har for nylig fået betydelig opmærksomhed (Figur 1B). Tab af Golgi-båndet er et fælles træk ved mange neurodegenerative sygdomme, herunder Alzheimers sygdom, Huntingtons sygdom, amyotrofisk lateralsklerose og Parkinsons sygdom (Gonatas et al., 2006; Haase og Rabouille, 2015; Rabouille og Haase, 2015; Rabouille og Haase, 2015; Sundaramoorthy et al., 2015). Det er meget sandsynligt, at forstyrrelserne i Golgi-arkitekturen i disse sygdomme bidrager til de patologiske processer. I de fleste tilfælde er den præcise morfologiske struktur af Golgi-fragmenterne i disse neurodegenerative sygdomme ikke blevet defineret. To eksperimentelle systemer har imidlertid for nylig undersøgt ændringer i Golgi-strukturen og neuronal degeneration i detaljer. For det første blev det påvist, at betinget knock out af GM130, et strukturelt golgin, som regulerer Golgi-båndet, i centralnervesystemet forårsager Golgi-fragmentering, atrofi af dendritter og neuronal degeneration hos mus (Liu et al., 2017). EM-analyse af GM130 KO Purkinjeceller viste en reduktion af Golgi cisternelængde og stabling, og desuden var de typiske Golgi dendritforposter fraværende i disse GM130 -/- Purkinjeceller (Liu et al., 2017). I en anden undersøgelse afslørede hippocampale neuroner fra mus transgene for den svenske mutation af amyloid precursor protein (APP) og en mutant presenilin 1-underenhed af γ-sekretase, hvor begge mutationer er forbundet med tidligt indsættende Alzheimers sygdom, omfattende Golgi-fragmentering ved optisk mikroskopi (Joshi et al., 2014). Kvantitativ EM af neuroner i disse mus viste en reduktion i antallet og længden af cisternerne i stakkene sammenlignet med neuroner fra vildtype-mus. Desuden var cisternerne opsvulmede. Ændringerne i Golgi-morfologien i disse primære neuroner blev vist at være direkte forbundet med det forhøjede niveau af amyloid-β-produktion. Primære neuroner behandlet med syntetisk amyloid-β viste også en lignende fragmentering af Golgi samt en stigning i tubulovesikulære strukturer forbundet med Golgi-cisternerne sammenlignet med ubehandlede celler. Tabet af Golgi-båndet skyldtes fosforylering af GRASP65, et Golgi-strukturprotein, som spiller en nøglerolle i adskillelsen af Golgi-båndet og -stakkene i mitose (Joshi et al., 2014). Sammenligning af disse to ovennævnte undersøgelser er informativ, da de veje, der formidler ændringerne i Golgi-morfologi, er forskellige i hvert tilfælde, hvilket fører til forskelle i morfologien af Golgi-“fragmenterne”. Det skal overvejes, hvordan disse forskellige veje, der påvirker Golgi-morfologien, påvirker downstream-reaktionerne.
mTOR-signalering
Vores laboratorium har etableret en eksperimentel tilgang til at forstyrre balancen mellem Golgi-båndet og Golgi-mini-stakkene ved at modulere dosis af GCC88, en golgin, der er placeret ved TGN. Denne strategi gjorde det muligt at etablere en stabil cellelinje, HeLa-B6, som mangler et Golgi-bånd. Vi har vist, at GCC88 regulerer balancen mellem Golgi-bånd og mini-stacks ved en aktinafhængig proces (Makhoul et al., 2019) og identificerede intersectin-1 (ITSN1), en guaninnukleotidudvekslingsfaktor for cdc42 (Hussain et al., 2001), som en interaktor af GCC88, der er ansvarlig for tabet af Golgi-båndet (Makhoul et al., 2019). Analyser af HeLa B6-celler, som mangler et Golgi-bånd, viste reduceret mTOR-aktivitet og en associeret stigning i autofagosombiogenese (Gosavi et al., 2018). mTOR er en af de vigtigste signalveje i eukaryote celler og kendt for at være en negativ regulator af autofagi (Wullschleger et al., 2006). Derfor har balancen mellem Golgi-stakke og Golgi-bånd en direkte effekt på mTORC1-banen. Brugen af cis- og trans-rumsspecifikke markører og EM-tomografi var afgørende for at afsløre de morfologiske ændringer i Golgi-båndet.
Inflammation
Inflammasomer i det medfødte immunsystem fungerer som et stillads for caspase 1-afhængig aktivering af proinflammatoriske cytokiner (Broz og Dixit, 2016). NLRP3 (nucleotide-binding domain, leucine-rich-containing family, pyrin domain-containing-3) er et alsidigt inflammasom, som kan aktiveres af en række mikrobielle og ikke-mikrobielle stimuli, hvilket resulterer i sekretion af pro-inflammatoriske cytokiner interleukin 1β (IL-1β) og interleukin 18 (IL-18) og programmeret celledød ved pyroptose. En nylig undersøgelse har vist, at aktiveringen af det cytosoliske NLRP3 ved stimuli involverer rekruttering af NLRP3 til spredte TGN-membraner for at lette NLRP3-saffold-samlingen (Chen og Chen, 2018). Den spredte TGN, men ikke de underliggende andre kompartmenter, rekrutterer specifikt NLRP3, via PI4P, til at samle downstream adaptorkomplekset ASC, som gennemgår polymerisering i den perikonukleære region før rekruttering af caspase-1 for at aktivere downstream-signalvejen (Chen og Chen, 2018). Derfor viser dette vigtige fund, at modulering af arkitekturen af TGN selektivt er kritisk i aktiveringen af denne vej. Brugen af kompartmentspecifikke markører (TGN og cis-Golgi) sammen med optisk og EM-analyse var afgørende for at afsløre de morfologiske ændringer i Golgi-båndet.
Sluttende bemærkninger
Den præcise arkitektur af Golgi-morfologien er defineret af den optiske mikroskopi med høj opløsning og af EM. Inddragelse af disse oplysninger i fremtidige undersøgelser på området vil give en betydelig indsigt i Golgis dynamik, vejene for forstyrrelse af båndstrukturen og de funktionelle konsekvenser forbundet med disse forskellige veje.
Sammenfattende foreslår vi, at brugen af udtrykket “fragmenteret Golgi” er utilstrækkelig til at beskrive Golgi-strukturer forbundet med mange behandlinger og tilstande, og forskellene i disse Golgi-strukturer er sandsynligvis relevante rent fysiologisk. I betragtning af Golgi-apparatets dynamiske karakter er det muligt, at der kan være en balance mellem forskellige morfologiske tilstande af Golgi på et givet tidspunkt, dvs. mini-stacks og båndstrukturer. Forståelse af den dynamiske balance mellem de forskellige Golgi-morfologier i molekylære detaljer er afgørende for en fuld forståelse af denne organel under normale celleprocesser og også under patologiske forhold. Det vil være fascinerende at se, hvad der udfolder sig, efterhånden som vi lærer mere om denne komplekse organelles celleafsøgende funktioner og forholdet mellem disse funktioner og Golgi-strukturer.
Datatilgængelighed
Alle datasæt, der er analyseret til denne undersøgelse, er inkluderet i manuskriptet og i de supplerende filer.
Author Contributions
Alle forfattere på listen har ydet et væsentligt, direkte og intellektuelt bidrag til arbejdet og har godkendt det til offentliggørelse.
Funding
Dette arbejde blev støttet af midler fra Australian Research Council (DP160102394).
Interessekonflikter
Forfatterne erklærer, at forskningen blev udført uden kommercielle eller finansielle forbindelser, der kunne opfattes som en potentiel interessekonflikt.
Abkortninger
GM130, cis-Golgi matrixprotein; GMAP210, Golgi microtubule associated protein; GOLPH3, Golgi phosphoprotein 3; GRASP, Golgi reassembly stacking protein; MTOC, microtubule organizing center; mTOR, mechanistic target of rapamycin; PI4P, phosphatidylinositol-4-phosphat; TGN, trans-Golgi network.
Abdel Rahman, A. M., Ryczko, M., Nakano, M., Pawling, J., Rodrigues, T., Johswich, A., et al. (2015). Golgi N-glykanforgrenende N-acetylglucosaminyltransferaser I, V og VI fremmer næringsstofoptagelse og metabolisme. Glycobiology 25, 225-240. doi: 10.1093/glycob/cwu105
PubMed Abstract | CrossRef Full Text | Google Scholar
Blackburn, J. B., og Lupashin, V. V. (2016). Oprettelse af knockouts af konserverede oligomeriske golgi-kompleksunderenheder ved hjælp af CRISPR-medieret genredigering parret med en udvælgelsesstrategi baseret på glykosyleringsdefekter, der er forbundet med nedsat COG-kompleksfunktion. Metoder Mol. Biol. 1496, 145-161. doi: 10.1007/978-1-4939-6463-5_12
PubMed Abstract | CrossRef Full Text | Google Scholar
Boncompain, G., og Perez, F. (2013). De mange ruter af Golgi-afhængig trafikering. Histochem. Cell Biol. 140, 251-260. doi: 10.1007/s00418-013-1124-7
PubMed Abstract | CrossRef Full Text | Google Scholar
Broz, P., og Dixit, V. M. (2016). Inflammasomer: mekanisme for samling, regulering og signalering. Nat. Rev. Immunol. 16, 407-420. doi: 10.1038/nri.2016.58
PubMed Abstract | CrossRef Full Text | Google Scholar
Chen, J., og Chen, Z. J. (2018). PtdIns4P på spredt trans-Golgi-netværk medierer NLRP3 inflammasomaktivering. Nature 564, 71-76. doi: 10.1038/s41586-018-0761-3
PubMed Abstract | CrossRef Full Text | Google Scholar
Chia, J., Goh, G., Racine, V., Ng, S., Kumar, P., og Bard, F. (2012). RNAi-screening afslører et stort signalnetværk, der kontrollerer Golgi-apparatet i menneskelige celler. Mol. Syst. Biol. 8:629. doi: 10.1038/msb.2012.59
PubMed Abstract | CrossRef Full Text | Google Scholar
Corda, D., Barretta, M. L., Cervigni, R. I., og Colanzi, A. (2012). Golgi-kompleksfragmentering i G2/M-overgangen: et organelbaseret cellecyklus-kontrolpunkt. IUBMB Life 64, 661-670. doi: 10.1002/iub.1054
PubMed Abstract | CrossRef Full Text | Google Scholar
De Matteis, M. A., Mironov, A. A. A., og Beznoussenko, G. V. (2008). Golgi-båndet og Golgins funktion. New York, NY: Springer-Verlag/Wein.
Google Scholar
Dippold, H. C., Ng, M. M. M., Farber-Katz, S. E., Lee, S. K., Kerr, M. L., Peterman, M. C., et al. (2009). GOLPH3 bygger bro mellem phosphatidylinositol-4-fosfat og actomyosin for at strække og forme Golgi for at fremme knopdannelse. Cell 139, 337-351. doi: 10.1016/j.cell.2009.07.052
PubMed Abstract | CrossRef Full Text | Google Scholar
Efimov, A., Kharitonov, A., Efimova, N., Loncarek, J., Miller, P. M., Andreyeva, N., et al. (2007). Asymmetrisk CLASP-afhængig kernedannelse af ikke-centrosomale mikrotubuli ved trans-Golgi-netværket. Dev. Cell 12, 917-930. doi: 10.1016/j.devcel.2007.04.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Farber-Katz, S. E., Dippold, H. C., Buschman, M. D., Peterman, M. C., Xing, M., Noakes, C. J., et al. (2014). DNA-skader udløser Golgi-spredning via DNA-PK og GOLPH3. Cell 156, 413-427. doi: 10.1016/j.cell.2013.12.023
PubMed Abstract | CrossRef Full Text | Google Scholar
Farhan, H., og Rabouille, C. (2011). Signalering til og fra den sekretoriske vej. J. Cell Sci. 124, 171-180. doi: 10.1242/jcs.076455
PubMed Abstract | CrossRef Full Text | Google Scholar
Farhan, H., Wendeler, M. W., Mitrovic, S., Fava, E., Silberberg, Y., Sharan, R., et al. (2010). MAPK-signalering til den tidlige sekretoriske vej afsløret ved funktionel screening af kinase/fosfatase. J. Cell Biol. 189, 997-1011. doi: 10.1083/jcb.200912082
PubMed Abstract | CrossRef Full Text | Google Scholar
Ferraro, F., Kriston-Vizi, J., Metcalf, D. J., Martin-Martin, B., Freeman, J., Burden, J. J., et al. (2014). En to-niveau Golgi-baseret kontrol af organelstørrelse underbygger endothelcellernes funktionelle plasticitet. Dev. Cell 29, 292-304. doi: 10.1016/j.devcel.2014.03.021
PubMed Abstract | CrossRef Full Text | Google Scholar
Gonatas, N. K., Stieber, A., og Gonatas, J. O. (2006). Fragmentering af Golgi-apparatet i neurodegenerative sygdomme og celledød. J. Neurol. Sci. 246, 21-30. doi: 10.1016/j.jns.2006.01.019
PubMed Abstract | CrossRef Full Text | Google Scholar
Gosavi, P., og Gleeson, P. A. (2017). Funktionen af Golgi-båndstrukturen- Et varigt myterium udfoldes! Bioessays 39:1700063. doi: 10.1002/bies.201700063
PubMed Abstract | CrossRef Full Text | Google Scholar
Gosavi, P., Houghton, F. J., Mcmillan, P. J., Hanssen, E., og Gleeson, P. A. (2018). Golgi-båndet i pattedyrceller regulerer autofagi negativt ved at modulere mTOR-aktivitet. J. Cell Sci. 131:jcs211987. doi: 10.1242/jcs.211987
PubMed Abstract | CrossRef Full Text | Google Scholar
Gunn, P. A., Gliddon, B. L., Londrigan, S. L., Lew, A. M., Van Driel, I. R., og Gleeson, P. A. (2011). Golgi-apparatet i de endomembranrige gastriske parietalceller eksisterer som funktionelle stabile mini-stakke spredt ud over hele cytoplasmaet. Biol. Cell 103, 559-572. doi: 10.1042/BC20110074
PubMed Abstract | CrossRef Full Text | Google Scholar
Haase, G., og Rabouille, C. (2015). Golgi-fragmentering i ALS-motorneuroner. Nye mekanismer, der er rettet mod mikrotubuli, tethers og transportvesikler. Front. Neurosci. 9:448. doi: 10.3389/fnins.2015.00448
PubMed Abstract | CrossRef Full Text | Google Scholar
Hussain, N. K., Jenna, S., Glogauer, M., Quinn, C. C., Wasiak, S., Guipponi, M., et al. (2001). Endocytisk protein intersectin-l regulerer aktin samling via Cdc42 og N-WASP. Nat. Cell Biol. 3, 927-932. doi: 10.1038/ncbb1001-927
PubMed Abstract | CrossRef Full Text | Google Scholar
Joshi, G., Chi, Y., Huang, Z., og Wang, Y. (2014). Abeta-induceret Golgi-fragmentering i Alzheimers sygdom øger Abeta-produktionen. Proc. Natl. Acad. Sci. U.S.A. 111, E1230-E1239. doi: 10.1073/pnas.1320192111
PubMed Abstract | CrossRef Full Text | Google Scholar
Kreft, M. E., Di Giandomenico, D., Beznoussenko, G. V., Resnik, N., Mironov, A. A. og Jezernik, K. (2010). Golgiapparatfragmentering som en mekanisme, der er ansvarlig for ensartet levering af uroplakiner til den apikale plasmamembran i uroepithelceller. Biol. Cell 102, 593-607. doi: 10.1042/bc20100024
PubMed Abstract | CrossRef Full Text | Google Scholar
Kupfer, A., Dennert, G., og Singer, S. J. (1983). Polarisation af Golgi-apparatet og det mikrotubuli-organiserende center inden for klonede naturlige dræberceller bundet til deres mål. Proc. Natl. Acad. Sci. U.S.A. 80, 7224-7228. doi: 10.1073/pnas.80.23.7224
PubMed Abstract | CrossRef Full Text | Google Scholar
Lamb, C. A., Yoshimori, T., og Tooze, S. A. (2013). Autofagosomet: Oprindelse ukendt, biogenese kompleks. Nat. Rev. Mol. Cell Biol. 14, 759-774. doi: 10.1038/nrm3696
PubMed Abstract | CrossRef Full Text | Google Scholar
Lasiecka, Z. M., og Winckler, B. (2011). Mekanismer for polariseret membrantrafik i neuroner – med fokus på endosomer. Mol. Cell. Neurosci. 48, 278-287. doi: 10.1016/j.mcn.2011.06.013
PubMed Abstract | CrossRef Full Text | Google Scholar
Lavieu, G., Dunlop, M. H., Lerich, A., Zheng, H., Bottanelli, F., og Rothman, J. E. (2014). Golgi-båndstrukturen letter anterograd transport af store ladninger. Mol. Biol. Cell 25, 3028-3036. doi: 10.1091/mbc.E14-04-093
PubMed Abstract | CrossRef Full Text | Google Scholar
Lazaro-Dieguez, F., Jimenez, N., Barth, H., Koster, A. J., Renau-Piqueras, J., Llopis, J. L. L., et al. (2006). Aktinfilamenter er involveret i vedligeholdelsen af Golgi-cisternernes morfologi og intra-Golgi pH. Cell Motil. Cytoskeleton 63, 778-791. doi: 10.1002/cm.20161
PubMed Abstract | CrossRef Full Text | Google Scholar
Liu, C., Mei, M., Li, Q., Roboti, P., Pang, Q., Ying, Z., et al. (2017). Tab af golgin GM130 forårsager Golgi-forstyrrelse, Purkinje-neuron tab og ataksi hos mus. Proc. Natl. Acad. Sci. U.S.A. 114, 346-351. doi: 10.1073/pnas.1608576114
PubMed Abstract | CrossRef Full Text | Google Scholar
Lu, Z., Joseph, D., Bugnard, E., Zaal, K. J., og Ralston, E. (2001). Golgi-kompleks reorganisering under muskeldifferentiering: visualisering i levende celler og mekanisme. Mol. Biol. Cell 12, 795-808. doi: 10.1091/mbc.12.4.795
PubMed Abstract | CrossRef Full Text | Google Scholar
Luini, A., og Parashuraman, S. (2016). Signalering ved Golgi: Afsporing og styring af membranfluxer. Curr. Opin. Cell Biol. 39, 37-42. doi: 10.1016/j.ceb.2016.01.014
PubMed Abstract | CrossRef Full Text | Google Scholar
Makhoul, C., Gosavi, P., Duffield, R., Delbridge, B., Williamson, N. A., og Gleeson, P. A. (2019). Intersectin-1 interagerer med golgin GCC88 for at koble aktinnetværket og Golgi-arkitekturen. Mol. Biol. Cell 30, 370-386. doi: 10.1091/mbc.E18-05-0313
PubMed Abstract | CrossRef Full Text | Google Scholar
Makhoul, C., Gosavi, P., og Gleeson, P. A. (2018). Golgi-arkitekturen og celleafsøgning. Biochem. Soc. Trans. 46, 1063-1072. doi: 10.1042/BST20180323
PubMed Abstract | CrossRef Full Text | Google Scholar
Mayinger, P. (2011). Signalering ved Golgi. Cold Spring Harb. Perspect. Biol. 3:a005314.
Google Scholar
Millarte, V., Boncompain, G., Tillmann, K., Perez, F., Sztul, E., og Farhan, H. (2015). Phospholipase C gamma1 regulerer tidlig sekretorisk trafikering og cellemigration via interaktion med p115. Mol. Biol. Cell 26, 2263-2278. doi: 10.1091/mbc.E15-03-0178
PubMed Abstract | CrossRef Full Text | Google Scholar
Millarte, V., og Farhan, H. (2012). Golgi i cellemigration: regulering ved signaltransduktion og dens implikationer for metastase af kræftceller. ScientificWorldJournal 2012:498278. doi: 10.1100/2012/498278
PubMed Abstract | CrossRef Full Text | Google Scholar
Miller, V. J., og Ungar, D. (2012). Re’COG’nition på Golgi. Traffic 13, 891-897. doi: 10.1111/j.1600-0854.2012.01338.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Petrosyan, A. (2015). Onco-Golgi: Er fragmentering en port til kræftprogression? Biochem. Mol. Biol. J. 1:16.
PubMed Abstract | Google Scholar
Puthenveedu, M. A., Bachert, C., Puri, S., Lanni, F., og Linstedt, A. D. (2006). GM130- og GRASP65-afhængig lateral cisternal fusion muliggør ensartet Golgi-enzymfordeling. Nat. Cell Biol. 8, 238-248. doi: 10.1038/ncb1366
PubMed Abstract | CrossRef Full Text | Google Scholar
Rabouille, C., og Haase, G. (2015). Editorial: Golgi-patologi i neurodegenerative sygdomme. Front. Neurosci. 9:489. doi: 10.3389/fnins.2015.00489
PubMed Abstract | CrossRef Full Text | Google Scholar
Rabouille, C., og Kondylis, V. (2007). Golgi ribbon unlinking: et organelbaseret G2/M checkpoint. Cell Cycle 6, 2723-2729. doi: 10.4161/cc.6.22.4896
PubMed Abstract | CrossRef Full Text | Google Scholar
Rambourg, A., og Clermont, Y. (1997). Tre-dimensionel struktur af Golgi-apparatet i pattedyrsceller. Basel: Birkhauser.
Google Scholar
Sanders, A. A., og Kaverina, I. (2015). Kernedannelse og dynamik af Golgi-afledte mikrotubuli. Front. Neurosci. 9:431. doi: 10.3389/fnins.2015.00431
CrossRef Full Text | Google Scholar
Sasaki, K., og Yoshida, H. (2015). Organelle autoregulering-stressreaktioner i ER, Golgi, mitokondrier og lysosom. J. Biochem. 157, 185-195. doi: 10.1093/jb/mvvv010
PubMed Abstract | CrossRef Full Text | Google Scholar
Scott, K. L., Kabbarah, O., Liang, M. C., Ivanova, E., Anagnostou, V., Wu, J., et al. (2009). GOLPH3 modulerer mTOR-signalering og rapamycinfølsomhed i kræft. Nature 459, 1085-1090. doi: 10.1038/nature08109
PubMed Abstract | CrossRef Full Text | Google Scholar
Serebrenik, Y. V., Hellerschmied, D., Toure, M., Lopez-Giraldez, F., Brookner, D., and Crews, C. M. (2018). Målrettet proteinudfoldning afdækker et Golgi-specifikt transkriptionelt stressrespons. Mol. Biol. Cell 29, 1284-1298. doi: 10.1091/mbc.E17-11-0693
PubMed Abstract | CrossRef Full Text | Google Scholar
Sundaramoorthy, V., Sultana, J. M., og Atkin, J. D. (2015). Golgi-fragmentering i amyotrofisk lateralsklerose, en oversigt over mulige udløsere og konsekvenser. Front. Neurosci. 9:400. doi: 10.3389/fnins.2015.00400
PubMed Abstract | CrossRef Full Text | Google Scholar
Wei, J. H., og Seemann, J. (2010). Optrævling af Golgi-båndet. Traffic 11, 1391-1400. doi: 10.1111/j.1600-0854.2010.01114.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Wei, J. H., og Seemann, J. (2017). Golgi-båndets disassemblering under mitose, differentiering og sygdomsprogression. Curr. Opin. Cell Biol. 47, 43-51. doi: 10.1016/j.ceb.2017.03.008
PubMed Abstract | CrossRef Full Text | Google Scholar
Wu, J., De Heus, C., Liu, Q., Bouchet, B. P., Noordstra, I., Jiang, K., et al. (2016). Molekylær vej for mikrotubuliorganisering ved Golgi-apparatet. Dev. Cell 39, 44-60. doi: 10.1016/j.devcel.2016.08.009
PubMed Abstract | CrossRef Full Text | Google Scholar
Wullschleger, S., Loewith, R., og Hall, M. N. (2006). TOR-signalering i vækst og metabolisme. Cell 124, 471-484.
Google Scholar
Yamamamoto, H., Kakuta, S., Watanabe, T. M., Kitamura, A., Sekito, T., Kondo-Kakuta, C., et al. (2012). Atg9 vesikler er en vigtig membrankilde under de tidlige trin af autofagosomdannelsen. J. Cell Biol. 198, 219-233. doi: 10.1083/jcb.201202061
PubMed Abstract | CrossRef Full Text | Google Scholar
Zappa, F., Failli, M., og De Matteis, M. A. (2018). Golgi-komplekset i sygdom og terapi. Curr. Opin. Cell Biol. 50, 102-116. doi: 10.1016/j.ceb.2018.03.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Leave a Reply