Frontiers in Marine Science
Introduction
Korale głębinowe są bardzo zróżnicowaną grupą organizmów morskich, a region Nowej Zelandii jest gorącym punktem dla korali głębinowych (Sánchez i Rowden, 2006; Cairns i in., 2009). W kilku badaniach przeanalizowano wpływ trałowania dennego na zespoły makrobezkręgowców głębinowych (w tym koralowce) w regionie Południowego Pacyfiku (Althaus i in., 2009; Clark i Rowden, 2009; Clark i in., 2010, 2019; Williams i in., 2010; Anderson i in., 2014). Wykazały one znaczące oddziaływanie trałowania dennego na wolno rosnące i wrażliwe kolonie i zbiorowiska koralowców głębinowych, których zdolność do regeneracji jest niewielka lub żadna. Przewiduje się, że wpływy środowiskowe przewidywane w związku ze zmianą klimatu, takie jak ocieplenie oceanu i zakwaszenie oceanu, również zagrażają zdrowiu koralowców głębinowych, przy czym niektóre siedliska stają się zbyt ciepłe, aby mogły się rozwijać w Anthony i Marshall (2009), lub obniżone pH i jony węglanowe, co prawdopodobnie wpłynie na koralowce głębinowe o szkieletach opartych na węglanach (Gammon i in., 2018). Aby lepiej informować przyszłe oceny ryzyka i zarządzanie zasobami morskimi, zrozumienie wieku i wzrostu koralowców głębinowych jest kluczowe dla określenia czasu regeneracji koralowców i zdolności do regeneracji po zakłóceniach antropogenicznych lub innych naturalnych uszkodzeniach.
W regionie Nowej Zelandii podjęto znaczną ilość prac dotyczących taksonomii koralowców głębinowych oraz rozmieszczenia różnych gatunków i ich preferencji środowiskowych, a także znaczenia koralowców głębinowych dla ekosystemów bentosowych. Przegląd naszego zrozumienia i wiedzy na temat koralowców głębokowodnych w regionie Nowej Zelandii został niedawno opublikowany (Tracey i Hjorvarsdottir, 2019). Przegląd ten nakreślił również główne luki w naszym zrozumieniu koralowców głębinowych, w tym ograniczone dane dotyczące wieku i wzrostu dla wielu rodzajów i gatunków. W tym badaniu oceniamy wiek i tempo wzrostu chronionego głębinowego koralowca czarnego Antipatharian Bathypathes patula. Wiek i tempo wzrostu tego koralowca nie były wcześniej badane w tym regionie. Wybór tego gatunku był oparty na liście priorytetów oceny ryzyka (Clark i in., 2014) i przeglądzie literatury (Tracey i in., 2018), w połączeniu z dostępnością próbek (lokalizacja i całkowita liczba), oraz innymi uzupełniającymi badaniami koralowców czarnych podejmowanymi w Nowej Zelandii (Hitt i in., 2020).
Tracey i in. (2018) nakreślili obecne metody, które są wykorzystywane do starzenia głębinowych gatunków koralowców. Omówiono zalety i wady każdej z metod. Główne metody, które zostały z powodzeniem zastosowane do pomiaru wieku i wzrostu koralowców głębinowych to (1) bezpośrednia obserwacja, taka jak pomiary in situ lub eksperymenty in-aquaria dotyczące wzrostu liniowego lub wydłużenia powierzchni; tempo dodawania polipów; oszacowanie tempa zwapnienia (np, przy użyciu techniki wagi wypornościowej), (2) wyliczanie stref wzrostu szkieletu lub pasm (sklerochronologia), oraz (3) analizy radiometryczne.
Bezpośrednie obserwacje i pomiary in situ lub in-aquaria są bardzo trudne do podjęcia w przypadku koralowców głębinowych, ponieważ trudno jest uzyskać żywe żywotne okazy i utrzymać je przy życiu, ponieważ ich wzrost trwa długo (Gammon i in., 2018; Tracey i in., 2018). Wyliczanie stref wzrostu, Sclerochronology, jest idealne dla głębinowych gatunków koralowców, które mają wysoki kontrast pomiędzy strefowymi wzorcami wzrostu obserwowanymi w przekrojach szkieletowych. Istotne jest jednak, aby zastosować niezależną metodę walidacji w celu sprawdzenia chronologicznej ważności zliczeń obserwowanych stref wzrostu (Andrews i in., 2002). Metoda uzyskiwania widocznych zliczeń stref wzrostu w strukturze szkieletowej została z powodzeniem zastosowana do określania danych dotyczących wieku i tempa wzrostu gorgoniańskich oktokorali, w tym 400-letnich koralowców bambusowych Keratoisis sp, 100-letniego seafana Primnoa spp. i 60-letniego koralowca bambusowego Lepidisis sp. (Thresher et al., 2004, 2007; Mortensen i Buhl-Mortensen, 2005; Sherwood et al., 2005; Tracey et al., 2007; Sherwood i Edinger, 2009). Korale czarne zostały również z powodzeniem zestarzone przy użyciu strefy wzrostu lub liczenia, z wiekiem od 150 lat do 480 lat dla Antipathes dendrochristos i Leiopathes glaberrima, odpowiednio (Love i in., 2007; Williams i in., 2007). W badaniach koralowca czarnego Stauropathes arctica uzyskano liczebność stref 55-58 lat (Sherwood i Edinger, 2009), przy czym autorzy zaznaczają, że datowanie radiowęglowe zawęziło ten wiek do 55 i 82 lat. Tempo wzrostu koralowców czarnych poprzez wyliczanie stref wzrostu ujawnia niski wzrost radialny od 8 do 140 μm/rok (Love i in., 2007; Prouty i in., 2011).
Istnieją potencjalne ograniczenia metody wyliczania stref wzrostu (patrz podsumowanie w Tracey i in., 2018). Wiele koralowców nie wykazuje wyraźnej regularnej struktury strefowej w całym swoim szkielecie, lub wyraźna struktura strefowa może być widoczna tylko w porcjach szkieletu. Nawet jeśli jest obecna, struktura strefowa może być zmienna lub wykazywać różne możliwości interpretacji. Na przykład liczebność podstaw i wierzchołków bambusowych gatunków oktokoralowców Keratoisis sp. i Lepidisis sp. była wyższa niż wiek oszacowany metodą radiometryczną z użyciem ołowiu 210 (210Pb) (Tracey i in., 2007). Postawiono hipotezę, że strefy obserwowane za pomocą mikroskopu świetlnego mają okresowość dwuletnią, a strefy obserwowane za pomocą SEM w węzłach mogą reprezentować wydarzenie środowiskowe, takie jak okresowość księżycowa. Cykle księżycowe mogą być związane z silnymi pływami, które mogą zwiększać dostępność pokarmu dla bezkręgowych organizmów żywiących się filtrem, powodując impulsy wzrostu somatycznego. Gorgoniańskie koralowce bąbelkowe (np. Paragorgia spp.) i czerwone koralowce szlachetne (np. Corallium sp.) nie zostały poddane udanemu procesowi starzenia przy użyciu liczenia strefowego ze względu na z natury skomplikowane lub słabo zdefiniowane wzory prążkowania (Griffin i Druffel, 1989; Andrews i in, 2005).
Najpowszechniejszą metodą radiometryczną stosowaną w przypadku koralowców głębinowych jest datowanie izotopem radiowęglowym (14C) (Druffel i in., 1995; Sherwood i in., 2005; Tracey i in., 2018). Metoda ta opiera się na fakcie, że bardzo mała ilość naturalnego węgla w atmosferze jest radioaktywna w postaci 14C i jest włączana do szkieletu koralowca, gdy tworzy on swój węglan wapnia lub białko i chitynę (Adkins i in., 2002; Tracey i in., 2003; Consalvey i in., 2006). Ponieważ okres połowicznego rozpadu radiowęglanu wynosi 5 730 lat, metoda ta, choć kosztowna, może być stosowana do wiarygodnego określania wieku okazów sprzed mniej niż 50 000 lat; po przekroczeniu tego okresu aktywność 14C staje się zbyt niska, by ją wykryć (Coleman, 1991). W latach 1950-60 nastąpił gwałtowny wzrost atmosferycznego 14C w wyniku testowania urządzeń jądrowych; ten „bombowy węgiel” może być również wykorzystywany jako narzędzie do kalibracji wieku, zapewniając punkt odniesienia dla nowszych próbek (Coleman, 1991; Druffel et al., 1995; Tracey et al., 2003; Roark et al., 2009; Sherwood i Edinger, 2009).
Radiocarbon dating has been successfully used on several coral species, including stony corals, black corals and gorgonian octocorals (Druffel et al., 1990; Roark et al., 2006; Carreiro-Silva et al., 2013; Prouty et al., 2016; Hitt et al., 2020). Metoda ta ma jednak również swoje ograniczenia. W wyniku cyrkulacji oceanicznej wiek radiowęglowy wody w oceanach jest znacznie starszy niż w atmosferze, tzw. „morski wiek zbiornikowy” (Tracey i in., 2003; Consalvey i in., 2006; Roark i in., 2006; Noé i in., 2008). Błędy w wieku zbiornikowym mogą mieć znaczący wpływ na szacunki wieku koralowców. Korygowanie wieku zbiorników wymaga wiedzy o otaczających wodach i zakłada stałe stosunki radiowęglowe w wodzie; jednak poziomy te mogą być zmienione przez mieszanie w oceanie, lokalne zdarzenia upwellingowe i zakwity produktywności, z których wszystkie mogą zmienić transformacje cyklu węglowego i skład 14C w szkielecie koralowców (Tracey i in., 2018).
Based on the Tracey et al. (2018) review we chose to use two ageing methods for B. patula; counts of skeletal growth bands and the radiometric method of radiocarbon dating (14C). Zastosowanie tych metod umożliwia porównania z innymi wcześniejszymi badaniami starzenia koralowców zarówno w Nowej Zelandii, jak i na świecie. Przedstawiono dobór próbek, metody mikro frezowania materiału i sekcjonowania oraz interpretację wyników liczenia stref i datowania radiowęglowego. Podano charakterystykę wieku i wzrostu korala czarnego B. patula i jednej kolonii Bathypathes cf. conferta (początkowo zakładanej jako B. patula).
Materiały i metody
Wybór próbek
Próbki użyte w tym badaniu pochodziły z zachowanych okazów złożonych w nowozelandzkim National Institute of Water and Atmospheric Research (NIWA), National Invertebrate Collection (NIC).
Próbki wybrano z dwóch regionów; Bay of Plenty (północno-wschodnia część Wyspy Północnej Nowej Zelandii) i Chatham Rise (wschodnia część Nowej Zelandii), w celu porównania wyników i oceny zmienności tempa wzrostu pomiędzy różnymi obszarami, a także z innymi głębinowymi rodzajami czarnych koralowców, w szczególności Antipathella, Leiopathes i Antipathes, z tych samych regionów (Hitt i in., 2020).
Prądy oceanograficzne wpływają na koralowce głębinowe, zapewniając stały dopływ pokarmu i rozprzestrzenianie się larw. W konsekwencji, prądy te wpływają również na dostępność ofiar i rozmieszczenie koralowców głębinowych. Podczas gdy na koralowce głębinowe największy wpływ mają prądy wody przydennej, które przepływają nad nimi i wpływają na dystrybucję osadów, oddziałują na nie również prądy wody powierzchniowej, które wpływają na produktywność pierwotną, a tym samym na dostawy pokarmu, które spływają do koralowców głębinowych na dnie morza. Lądolód Nowej Zelandii leży na skrzyżowaniu ciepłych, ubogich w składniki odżywcze, subtropikalnych wód powierzchniowych napływających z północy i chłodnych, bogatych w składniki odżywcze, subantarktycznych wód powierzchniowych napływających z południa. Obecne rozumienie mas wodnych i prądów oceanicznych w tym regionie na powierzchni, termoklinie, wodach pośrednich i głębokich zostało ostatnio przejrzane przez Chiswell et al. (2015). Ustawienie oceanograficzne dla koralowców z tego badania jest następujące.
Zatoka Plenty jest skąpana w ciepłych, ubogich w składniki odżywcze, subtropikalnych wodach powierzchniowych, które pochodzą z Prądu Wschodnioaustralijskiego i wpływają z zachodu, tworząc Prąd WschodnioAucklandzki. Chatham Rise jest miejscem spotkania ciepłych, o niskiej zawartości składników odżywczych, wód subtropikalnych (STW) i chłodnych, o wysokiej zawartości składników odżywczych, wód subantarktycznych (SAW) pochodzących z Oceanu Południowego jest Front Subtropikalny (STF). Mieszanie się mas wody STW i SAW na STF powoduje, że Chatham Rise jest strefą bardzo wysokiej produktywności pierwotnej dostarczającej znaczącego źródła pokarmu, a tym samym węgla dla tych koralowców, stężenia chlorofilu a wahają się od 0,3 do 0,48 mg/m3 w Chatham Rise i Bay of Plenty, w porównaniu do 0,03-0,3 mg/m3 w centralnej części Pacyfiku (Pinkerton, 2016). Na głębokościach 500-1500 m do tego regionu wpływają z północy i południa chłodne (3,5-10°C), o niskim zasoleniu (34,4-34,6) i wysokiej zawartości tlenu (200-250 μmol/kg) oraz składników odżywczych (azotany 15-36 μmol/kg) antarktyczne wody pośrednie (AAIW) (Chiswell i in., 2015).
Korale z Chatham Rise zostały wybrane, ponieważ istnieją współczesne dane dotyczące wieku rezerwuaru 14C z tego obszaru (Sikes i in., 2008). Znajomość lokalnego wieku rezerwuarowego 14C umożliwia bardziej precyzyjne oszacowanie wieku radiowęglowego koralowców.
Próbki zostały wybrane z istniejących okazów zebranych przez Fisheries New Zealand (FNZ), obserwatorów Ministry for Primary Industries (MPI) oraz badaczy NIWA i są przechowywane w (NIC). Wykres danych dotyczących rozmieszczenia B. patula pomógł w podjęciu decyzji o wyborze miejsca kolonii i liczbie próbek (Rysunek 1). Dwa okazy z Bay of Plenty (NIWA24190 i NIWA85940) zostały wybrane do datowania radiowęglowego, a cienkie przekroje do liczenia stref. Pozostałe okazy wybrano z Chatham Rise.
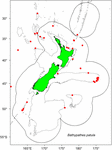
Figura 1. Lokalizacje, w których pobierano próbki Bathypathes patula w rejonie Nowej Zelandii. Czerwone kropki oznaczają indywidualne miejsca pobrania próbek koralowców z okazów B. patula przechowywanych w National Invertebrate Collection of New Zealand (NIC). Szare linie oznaczają kontur głębokości wody 200 m. Czarna linia oznacza zasięg wyłącznej strefy ekonomicznej Nowej Zelandii (EEZ).
Pod koniec 2018 roku wizytujący specjalista w dziedzinie taksonomii czarnych koralowców (Jeremy Horowitz, James Cook University) dokonał przeglądu identyfikacji kilku próbek w NIC. Podczas tego procesu jedna z kolonii czarnych koralowców wybranych do tych badań szacowania wieku i zakładana jako Bathypathes patula, otrzymała zrewidowaną identyfikację. Koralowiec czarny NIWA24190 został zidentyfikowany jako Bathypathes cf. conferta. W niniejszym opracowaniu próbka jest określana jako Bathypathes cf. conferta (Rysunek 2), co jest najbardziej prawdopodobną prawidłową identyfikacją. Dalsza weryfikacja taksonomiczna tego okazu jest w toku (Opresko pers. comm). Mapa rozmieszczenia (Ryc. 1) może zawierać dodatkowe przykłady B. cf. conferta.

Figura 2. Bathypathes cf. conferta. NIWA24190. Okaz pokazujący, gdzie pobrano próbki do analizy. (A) to wyrastający wierzchołek, (B) przekrój podstawowy. Okaz z Bay of Plenty, Nowa Zelandia. Pasek skali to linijka 35 cm.
Przygotowanie materiału
Gdy odpowiednie próbki zostały zidentyfikowane, małe (mniej niż 10 mm odcinki) zostały wycięte z rosnących końcówek i części bazalnej głównej łodygi koralowców. Ponieważ wiele z próbek zostało złamanych podczas procesu zbierania, dodatkowe odcinki zostały pobrane wzdłuż głównej łodygi, tak aby można było uzyskać wiarygodne szacunki liniowego tempa wzrostu (Rysunki 2-5). Próbki z rosnących końcówek zostały następnie podzielone na dwa fragmenty, jeden do datowania radiowęglowego, drugi do przygotowania cienkich przekrojów w celu uzyskania liczby stref.

Rysunek 3. Bathypathes patula NIWA85940. Okaz pokazujący, gdzie pobrano próbki do analizy. (A) to rosnący wierzchołek, (C) sekcja podstawowa, a (B) to pośrednie miejsca pobrania próbek, ponieważ okaz był złamany podczas zbierania i powyżej tego miejsca może brakować nieznanej długości głównego rozgałęzionego pnia. Okaz z Bay of Plenty, Nowa Zelandia. Pasek skali to linijka 35 cm.

Rysunek 4. Bathypathes patula NIWA49468. Okaz pokazujący, gdzie pobrano próbki do analizy. (A) to rosnący wierzchołek, (D) przekrój podstawowy, a (B,C) to pośrednie miejsca pobrania próbek, ponieważ okaz był złamany podczas zbierania i może brakować nieznanej długości głównego pnia gałęzi pomiędzy tymi dwoma miejscami. Okaz z Chatham Rise, Nowa Zelandia. Pasek skali to linijka 35 cm.

Figure 5. Bathypathes patula NIWA47911. Okaz pokazujący, gdzie pobrano próbki do analizy. (A) to rosnący wierzchołek, (F) to przekrój podstawowy, a (B-E) to pośrednie miejsca pobrania próbek w poszukiwaniu sygnału bomby radiowęglowej. Okaz z Chatham Rise, Nowa Zelandia. Pasek skali to linijka 35 cm.
Mikrofrezowanie materiału
W poprzednich pracach zauważono, że sekcje czarnego koralowca są podatne na wciąganie żywicy przez ich porowatą matrycę poprzez działanie kapilarne i dlatego sekcje w tym badaniu nie mogły być połączone żywicą z płytą bazową do frezowania. Zamiast tego wyprodukowano uchwyt do przytrzymywania sekcji podczas procesu frezowania, eliminując w ten sposób ryzyko zanieczyszczenia żywicą próbek radiowęglowych.
Sproszkowane próbki radiowęglowe zostały wyekstrahowane z krawędzi i rdzenia sekcji bazowych, środkowych i wierzchołkowych przy użyciu mikromłynka New WaveTM z frezem trzpieniowym 0,5 mm Brassler H2.11.006 w NIWA. Próbki z krawędzi zawierały materiał z zewnętrznych 250 mikronów przekroju, próbki z rdzenia były uzyskane z materiału w promieniu 750 mikronów od primordium. Staraliśmy się uzyskać co najmniej 1.5 mg materiału z każdej próbki. Próbki zostały następnie zważone i przeanalizowane pod kątem radiowęglowym w Australian National University (ANU) Radiocarbon Laboratory.
Metoda cienkich przekrojów i protokoły liczenia stref
Sekcje łodyg koralowców zostały zatopione w przezroczystej, dwuczęściowej żywicy epoksydowej i przekrojone piłą diamentową w NIWA. Sekcje były polerowane z jednej strony i montowane polerowaną stroną w dół na szklanym szkiełku mikroskopowym, następnie szlifowane i polerowane aż do uzyskania optymalnej grubości do oglądania struktury strefy wzrostu przy użyciu światła przechodzącego. Optymalna grubość dla sekcji tego gatunku wynosiła około 250-300 μm i była zgodna ze standardową procedurą przygotowywania cienkich sekcji (Andrews et al., 2002; Mortensen i Buhl-Mortensen, 2005; Tracey et al., 2007). Liczenie okresowych pasm wzrostu lub przyrostów, zakładanych jako tworzące się corocznie, miało miejsce po przygotowaniu cienkich przekrojów w NIWA.
Aby wygenerować liczenie stref, preparaty cienkich przekrojów głównych odcinków gałęzi były oglądane pod mikroskopem złożonym w świetle przechodzącym. Jako pomoc w interpretacji stref, sekcje były również oglądane przy użyciu światła ultrafioletowego zgodnie z metodologią Sherwood i Edinger (2009). Tutaj zliczanie stref odbywało się przy użyciu transmitowanego oświetlenia jasnego pola, a oświetlenie ultrafioletowe było użyteczną pomocą w dalszym definiowaniu zaobserwowanej struktury stref.
Metoda datowania radiowęglowego
Przed analizami, próbki zostały oczyszczone (np. wypłukane kwasem) z czarnych skorup i aktywności endolitycznej, aby usunąć wszelkie potencjalne młodsze zanieczyszczenia 14C, które mogą zmienić wyniki (Adkins i in., 2002). Sproszkowane próbki koralowców poddano działaniu <1 ml 0,1 M HCL. Kwas został następnie odpipetowany, a próbki przepłukano trzykrotnie wodą milliQ. Próbka została następnie zamrożona suchym lodem i zliofilizowana. Około 1mg oczyszczonej próbki koralowca odważono do kwarcowej probówki o średnicy 6 mm OD, dodano ∼60 mg CuO i srebrny kubek. Probówki próżniowano do <3e-3 torr (<0.4 Pa) i uszczelniano za pomocą palnika. Następnie probówki były wypalane w temperaturze 900°C przez 6 h w celu wytworzenia CO2. Powstały CO2 oczyszczono i przekształcono w grafit w obecności wodoru z zasilanym Fe jako katalizatorem. Grafit był następnie mierzony na jednostopniowym akceleratorowym spektrometrze mas ANU (Fallon et al., 2010). Wszystkie dane zostały skorygowane przy użyciu on-line AMS δ13C, znormalizowane do Oxalic Acid I i odjęte od tła przy użyciu węgla wolnego od 14C, traktowanego w taki sam sposób jak próbki koralowców. Dane przedstawiono zgodnie z zaleceniami Stuivera i Polacha (1977).
Wyniki
Sekcje cienkie dla liczeń strefowych
Strefowe wzory strukturalne obserwowane na sekcjach szkieletowych dla tych gatunków są bardzo złożone i mogą być interpretowane na wiele sposobów. W przypadku liczenia wzrostu promieniowego, przygotowane przekroje były dokładnie obserwowane, a oś, która najlepiej wykazywała wyraźną strukturę strefową od primordium do zewnętrznej krawędzi, była następnie używana do uzyskania liczby stref. Wzrost promieniowy rozgałęzień u B. patula był w miarę regularny, nie zaobserwowano wyraźnej tendencji w odkładaniu się stref tworzących grubsze strefy po konkretnej stronie rozgałęzienia, co wcześniej obserwowano u innych gatunków koralowców czarnych. Promienie użyte do obliczeń tempa wzrostu były średnią minimalnych i maksymalnych promieni zaobserwowanych w przygotowanych przekrojach, co oznacza, że podane tempo wzrostu radialnego będzie średnim tempem wzrostu radialnego dla koralowca.
Na szkielecie koralowca występuje początkowo gruboziarnisty wzór pasmowy w dużej mierze zdefiniowany przez naprzemienne ciemniejsze i jaśniejsze strefy (Rysunek 6) przy obserwacji pod mniejszą mocą (10-20×). Przy bliższym badaniu pod wyższą mocą (100-200×), widoczny jest również dość regularny wzór pasmowania w drobnej skali (Rysunek 7). Zastosowaliśmy tu dwa protokoły odczytu – licząc zarówno grubo uformowane szerokie strefy, jak i cienkie, drobno uformowane strefy (Tabela 1). Wszystkie przedstawione tu zliczenia stref zostały wykonane przez jednego czytelnika (Marriott). W trakcie opracowywania protokołów interpretacji stref dokonano wielu odczytów, stosując różne iteracje opracowywanych protokołów. Liczby stref z dwóch przedstawionych tu protokołów zostały uzyskane z wielu odczytów w całej sekcji.

Rysunek 6. Przekrój bazalny koralowca NIWA47911 ukazujący interpretację gruboziarnistego bandingu zaobserwowanej struktury strefowej. Na okazie widoczne są 54 strefy zaznaczone czerwonymi kropkami. Wstawiony przerywany prostokąt pokazuje centralną część przekroju bazalnego obrazowanego na Rysunku 7.

Figura 7. Środkowa część przekroju bazalnego koralowca NIWA47911 przedstawiająca protokół interpretacji w drobnej skali zaobserwowanej struktury strefowej. Okaz wykazuje 42 strefy oznaczone czarnymi kropkami w tym wewnętrznym regionie przekroju; cały przekrój wykazał 211 stref.

Tabela 1. Liczba stref z cienkich przekrojów.
W najmłodszej części badanych okazów, w rejonie wierzchołka lub w jego pobliżu, liczba stref wynosiła od 4 do 7 (liczba stref grubych) i od 10 do 19 (liczba stref drobnych). W środkowym regionie głównej gałęzi liczebność wahała się od 55 do 60 (liczebność w strefie grubej) i od 370 do 374 (liczebność w strefie drobnej). W rejonie podstawy kolonii liczebność wahała się od 37 do 193 (liczebność strefy grubej) i od 78 do 481 (liczebność strefy drobnej). Złożoność wzorów zonacji podkreśliła potrzebę walidacji.
Analiza radiowęglowa
Próbki zmielone wymienione w Tabeli 2 zostały wysłane do ANU w celu datowania radiowęglowego. Wyniki z 17 próbek koralowców są przedstawione w wynikach radiowęglowych (Tabela 3). Trzy próbki (próbki 47911-F2; 24190-B1; 85940-B1) uległy zniszczeniu w laboratorium, a pozostały materiał był niewystarczający do wykonania duplikatu. Wyniki radiowęglowe z pozostałych koralowców zostały skalibrowane do wieku kalendarzowego przy użyciu OxCal4.3 (Bronk Ramsey, 2001), krzywej Marine13 i nowozelandzkiego lokalnego deltaR (radiocarbon reservoir offset) wynoszącego -18 ± 36 (Reimer i Reimer, 2001). Rozkłady przedziałów wiekowych dla próbek koralowców sprzed wybuchu bomby przedstawiono w Tabeli 3. Wszystkie okazy koralowców użyte w niniejszych badaniach były żywe w momencie schwytania, więc wszelkie podane szacunki wieku dotyczą wieku w momencie schwytania, a nie długości życia okazu. Przewidywana długość życia dla tego gatunku będzie przekraczać przedstawione tu szacunki wieku. Koralowiec 85940 miał około 380 ± 50 lat, koralowiec 49468 około 310 ± 70 lat, a koralowiec 47911 około 120 ± 50 lat (Tabela 3). Wszystkie próbki zewnętrznych krawędzi i końcówek koralowców zawierały wartości radiowęglowe F14C >1 (Tabela 3), z wyjątkiem 85940-B2 i 85940-C2, co wskazuje na to, że koralowce te były żywe i zawierały węgiel z wód powierzchniowych w czasie pobierania. Korale 85940-B2 i 85940-C2 miały na zewnętrznych krawędziach wartości radiowęglowe F14C <1 (Tabela 3) wskazujące, że część szkieletu nie dodawała aktywnie materiału. Zaobserwowano to wcześniej, gdzie bazowa część kolonii koralowej przestaje rosnąć, jednak górna część nadal rośnie (Komugabe-Dixson i in., 2016).
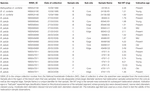
Tabela 2. Próbki pobrane do datowania radiowęglowego.

Tabela 3. Radiocarbon results.
Growth Rate Estimates
Oszacowania tempa wzrostu oparto na wynikach radiowęglowych, ponieważ liczenie stref było problematyczne i okazało się niepotwierdzone, więc nie mogło być wykorzystane do wygenerowania wiarygodnych oszacowań tempa wzrostu (patrz sekcja „Bathypathes patula Age and Growth Estimates” w celu dalszej dyskusji na temat liczenia stref). Wskaźniki wzrostu wyprowadzono na podstawie fizycznych odległości między punktami poboru próbek i skalibrowanych wieków radiowęglowych w danym miejscu poboru próbek. Ze względu na to, że niektóre próbki zawiodły podczas analizy radiowęglowej, można było wygenerować tylko ograniczony zestaw szacunków tempa wzrostu. Wskaźniki wzrostu radialnego, czyli tempo, w jakim gałąź kolonii zagęszcza się promieniście w czasie, zostały oszacowane poprzez podzielenie skalibrowanych lat kalendarzowych wzrostu pomiędzy rdzeniem a krawędzią miejsca pobrania próbki przez średni promień segmentu gałęzi. Wzrost radialny wahał się od 11,1 do 35,7 μm/rok. Liniowe tempo wzrostu, czyli tempo, w jakim gałązka kolonii wydłuża się wzdłużnie w czasie, zostało oszacowane poprzez podzielenie różnicy pomiędzy skalibrowanym wiekiem kalendarzowym próbek rdzenia przez liniową odległość pomiędzy dwoma miejscami poboru próbek. Uzyskano dwa liniowe tempa wzrostu, 5,2 i 9,6 mm/rok (Tabela 4).

Tabela 4. Bathypathes patula growth rate estimates.
Discussion
Bathypathes patula Age and Growth Estimates
Wyniki radiowęglowe z tej pracy pokazują, że B. patula jest gatunkiem długowiecznym, z wiekiem przekraczającym 385 lat. Wolne tempo wzrostu liniowego (5,2-9,6 mm/rok) i radialnego (11,1-35,7 μm/rok) uzyskano z danych dotyczących wieku radiowęglowego.
Żaden z protokołów liczenia stref nie korelował wiarygodnie z niezależnymi szacunkami wieku radiowęglowego. Protokół liczenia stref w drobnej skali wykazał najbliższą korelację z danymi dotyczącymi wieku 14C, zapewniając pewne ogólne wsparcie dla długowieczności przy porównywaniu liczenia stref z datami 14C. Oszacowania wieku między danymi dotyczącymi wieku radiowęglowego a danymi dotyczącymi liczenia stref w drobnej skali rutynowo wykazywały rozbieżności przekraczające sto lat w sekcjach oddziałów ze skalibrowanym wiekiem radiowęglowym 112-385 lat (Tabele 1, 3). Różnice w wieku szacowanym z porównywalnych próbek przy użyciu obu metod były ujemne i dodatnie; nie stwierdzono zaniżania ani zawyżania wieku przy użyciu liczenia strefowego o proporcjonalną wartość (Tab. 1). Oznacza to, że obserwowana struktura strefowa nie wykazuje rocznej, ani żadnej innej regularnej chronologicznej okresowości, a nasze protokoły liczenia stref nie mogą być stosowane do wiarygodnego generowania szacunków wieku dla B. patula.
Wyniki radiowęglowe z krawędzi okazu NIWA-85940 sugerują, że sekcja podstawowa (C2) głównego pnia przestała odkładać materiał szkieletowy na swojej zewnętrznej powierzchni 293 lata przed zbiorem, a sekcja środkowa (B2) przestała odkładać materiał szkieletowy 88 lat przed zbiorem. Stało się tak pomimo obserwacji, że te odcinki rozgałęzionego pnia bezpośrednio przylegały do bocznych pinnoli z żywymi polipami. Optyczny wygląd tych odcinków nie wskazywał na to, że na ich zewnętrznej krawędzi zaprzestano odkładania strefy. Powierzchnia gałązki koralowca nadal wydawała się być stosunkowo czysta, nie wykazywała żadnych oznak erozji powierzchniowej ani odbarwień, czego można by się spodziewać po gałązce koralowca, która nie akreuje już nowego materiału szkieletowego na swojej zewnętrznej powierzchni. Sugeruje to, że jeśli łodygi gałęzi mogą przestać tworzyć warstwy szkieletowe, podczas gdy końce gałęzi nadal rosną, wówczas protokoły metody liczenia strefowego nie mogą być wiarygodnie wykorzystane do generowania szacunków wieku z sekcji bazalnych dla tego gatunku koralowca.
W okazie NIWA47911 pobrano pięć próbek z rosnącej końcówki głównej gałęzi osiowej w kierunku podstawy koralowca. Próbki te były rozmieszczone w odległości 25 mm od siebie. Była to próba zlokalizowania pojawienia się bomby radiowęglowej w szkielecie koralowca. Oznaczałoby to, że liniowe tempo wzrostu mogłoby być dokładnie ocenione dla ostatnich rosnących końcówek gałęzi kolonii. Jeśli wzrost radiowęglowodoru bomby mógłby być dokładnie umiejscowiony, wtedy dałoby to również dokładny punkt odniesienia dla ustawienia poprawki 14C do zastosowania w próbkach radiowęglowodoru sprzed bomby dla tej kolonii i dla innych kolonii z tej samej masy wody. Niestety, wszystkie te próbki okazały się być post-bombowe, więc proces ten nie mógł być zakończony. Przyszłe pobieranie próbek dalej w dół tego pnia gałęzi byłoby przydatne do ustalenia tego punktu stycznego i współczesnego tempa wzrostu liniowego. Nasza praca mierzyła ogólne tempo wzrostu liniowego 5.2-9.6 mm/rok, zakładając najszybsze tempo wzrostu należałoby pobrać próbki 700 mm w dół głównego pnia gałęzi od rosnącego wierzchołka, aby mieć pewność zaobserwowania wzrostu radiowęglowodoru z bomby w szkielecie.
Badania Love et al. (2007) na gatunku czarnego korala Antipathes dendrochristos również podały dwie interpretacje struktury strefowej podobne do opisanych tutaj protokołów grubego i drobnego. Autorzy ci stwierdzili jednak, że grubiej zabarwiona struktura pasmowa korelowała z ich wynikami radiowęglowymi i 210Pb wskazującymi na pasmowość roczną (Love i in., 2007).
Pomiary radiowęglowe z obecnej pracy wykazały F14C >1 w próbkach z rosnących końcówek gałęzi i powierzchni głównej gałęzi, wskazując, że próbki te wykazywały współczesne poziomy radiowęglowe po wybuchu bomby. Obecność węgla po wybuchu bomby poniżej mieszanej warstwy oceanu wskazuje, że korale czarne B. patula wykorzystują niedawno wyeksportowane POC z wód powierzchniowych. Na tych głębokościach POC jest jedynym źródłem węgla oznaczonym przy pomocy bomby 14C. Rozpuszczony węgiel nieorganiczny (DIC) w wodzie otaczającej, w której żyją koralowce, ma znacznie niższe wartości 14C (Matsumoto, 2007; Alves i in., 2018). Wykrycie bomby 14C u koralowców, które rosną w wodach, które nie doświadczyły bomby 14C (tj. poprzez wentylację oceanu) wzmacnia, że czarne koralowce czerpią swój węgiel z POC pochodzącego z powierzchni (Roark i in., 2009; Sherwood i Edinger, 2009; Williams i Grottoli, 2010; Prouty i in., 2011). Oznacza to również, że nie musimy modelować efektu rezerwuaru głębokowodnego, jak ma to miejsce w przypadku koralowców kamiennych, które czerpią swój węgiel szkieletowy z DIC w wodach pośrednich, które je kąpią.
Porównania z ostatnimi nowozelandzkimi i globalnymi danymi o wieku czarnych koralowców
Oszacowania wieku i tempa wzrostu uzyskane radiowęglowo w tym projekcie są podobne do wyników kilku innych badań wieku i wzrostu, które zostały ostatnio podjęte dla kluczowych głębinowych czarnych koralowców (Antipatharia) w regionie Nowej Zelandii (Tabela 5). Te szacunki wieku i tempa wzrostu dla B. patula są podobne do innych rodzajów koralowców, takich jak kolonie Leiopathes, Antipathes i Antipathella, które żyją do 2600 lat i rosną w tempie 1-143 μm/rok (Hitt et al., 2020). Nasze wyniki dostarczają dalszych dowodów na to, że nowozelandzkie korale czarne są jednymi z najwolniej rosnących zwierząt morskich w regionie, przy czym niektóre gatunki wykazują ekstremalną długowieczność.

Tabela 5. Radiocarbon age estimation studies on New Zealand black corals (Antipatharia).
Wyniki badań B. patula i innych czarnych koralowców, które poddano starzeniu w Nowej Zelandii, nie wykazują wyraźnych trendów w tempie wzrostu lub długowieczności w odniesieniu do mas wodnych i głębokości, na których koralowce te występują. Próbki z Chatham Rise i Bay of Plenty pochodziły głównie z podobnych głębokości, od 750 do <1269 m. Głębszy okaz B. patula z Chatham Rise pochodził z głębokości pomiędzy 1120 a 1269 m (Tabele 1, 5). Tempo wzrostu radialnego naszych próbek z Chatham Rise (NIWA-49468 i NIWA47911) na poziomie 20-36 μm/rok było wyższe niż okazu z Zatoki Plenty (NIWA-85940) na poziomie 11 μm/rok. Ponieważ udało się uzyskać tylko trzy wskaźniki wzrostu dla próbek B. patula z tych dwóch regionów, nie można przyjąć żadnych założeń dotyczących regionalnych różnic w tempie wzrostu. Spektrum danych dotyczących tempa wzrostu koralowców czarnych z Chatham Rise pokazuje, że próbka Antipathes sp. miała najszybsze zaobserwowane tempo wzrostu, 91-143 μm/rok, jak również jedno z najwolniejszych, 5-15 μm/rok dla okazu Leiopathes secunda. Okazy korala czarnego z Zatoki Plenty: Bathypathes patula, Leiopathes secunda i Leiopathes sp. wykazują średnie lub niskie tempo wzrostu radialnego, 5-40 μm/rok. W przeciwieństwie do nich, płytkowodne czarne korale Fiordland Antipathella fiordensis mają stosunkowo wysokie tempo wzrostu promieniowego (50-57 μm/rok), ale nie jest jasne, czy jest to funkcja gatunku, czy warunków środowiskowych fiordów, czy też mieszanka obu.
Badania te zgadzają się również z globalnymi badaniami, które wykazały, że czarne korale są jednymi z najwolniej rosnących koralowców głębinowych. Sherwood i Edinger (2009) podają szacunkowy wiek czarnego korala Stauropathes arctica do 82 lat kalendarzowych. Prouty et al. (2011) wykorzystali radiowęglowodory do oszacowania wieku żywych kolonii Leiopathes sp. do 2100 lat. Carreiro-Silva et al. (2013) postarzyli azorską kolonię Leiopathes sp. na 2320 lat. Okaz Leiopathes annosahas z Hawajów wykazywał potencjał do osiągnięcia długości życia przekraczającej 4000 lat (Roark et al., 2009).
Te nowe wskaźniki wieku i wzrostu dla delikatnego głębinowego korala czarnego B. patula podkreślają, że jest to gatunek długowieczny, wykazany w tym badaniu do osiągnięcia wieku przekraczającego 385 lat, i wykazuje powolne liniowe i radialne tempo wzrostu. Wyniki wskazują na prawdopodobnie niską odporność i powolny czas regeneracji po zaburzeniu/usunięciu. Badania te zostaną wykorzystane w przyszłych ocenach ryzyka i strategiach zarządzania w nowozelandzkich ramach zarządzania koralowcami (Freeman i Cryer, 2019). Powolny wzrost i ekstremalna długowieczność dodatkowo podkreślają, że korale te będą bardzo podatne na podmorskie zakłócenia, takie jak rybołówstwo i górnictwo, i mogą potrzebować wieków, aby odzyskać siły po wpływach antropogenicznych (Hitt i in., 2020).
Podsumowanie
Wiek i tempo wzrostu głębinowych próbek czarnego korala B. patula z dwóch regionów Nowej Zelandii, Bay of Plenty i Chatham Rise, zostały określone przy użyciu dwóch różnych metod; liczenia pierścieni wzrostu (sklerochronologia) i datowania radiowęglowego. Do liczenia pierścieni wzrostu zastosowano dwa różne protokoły, wykorzystujące ciemne i jasne warstwy wzrostu. Dało to zróżnicowane wyniki, które były również niespójne z wiekami radiowęglowymi kilku z tych koralowców. Wieki radiowęglowe z B. patula wskazują, że ten gatunek koralowca żyje długo, z wiekiem ponad 385 lat, z powolnym liniowym (5,2-9,6 mm/rok) i radialnym (11,1-35,7 μm/rok) tempem wzrostu. Wyniki te są porównywalne z wiekiem i tempem wzrostu innych rodzajów koralowców czarnych z tych samych regionów Nowej Zelandii i z całego świata. Wiek i tempo wzrostu gatunków głębinowych są krytyczne dla określenia potencjalnego ryzyka tych organizmów na zaburzenia antropogeniczne. Wyniki te będą wykorzystane do informowania przyszłych ocen ryzyka i strategii zarządzania w nowozelandzkich ramach zarządzania koralowcami.
Oświadczenie o dostępności danych
Wszystkie zestawy danych wygenerowane dla tego badania są zawarte w artykule/materiale uzupełniające.
Wkład autorów
PM, DT, HB, i NH przyczynili się do koncepcji i projektu badania. PM wykonał wszystkie czynności związane z przygotowaniem próbek, analizą obrazu przekroju, opracowaniem protokołu cienkiego przekroju i liczeniem stref oraz mikromieleniem próbek radiowęglowych. NH oznaczył próbki radiowęglowe. SF przygotował poprawione oszacowania wieku na podstawie surowych danych radiowęglowych i napisał część manuskryptu. PM i DT napisali pierwszy projekt manuskryptu. All authors contributed to the manuscript revision.
Funding
This research was funded from Year Two of the Conservation Services Programme (CSP), Department of Conservation (DOC) Project POP2017-07 Objective to „Develop a methodology to determine the age and growth characteristics of key high-risk New Zealand deep-sea (cold-water) coral species.” Finansowanie opłaty za publikację w tym czasopiśmie zostało zapewnione przez NIWA, Nowa Zelandia.
Konflikt interesów
Autorzy deklarują, że badania zostały przeprowadzone przy braku jakichkolwiek komercyjnych lub finansowych relacji, które mogłyby być interpretowane jako potencjalny konflikt interesów.
Podziękowania
Dziękujemy Conservation Services Programme Group, Department of Conservation New Zealand, szczególnie Ianowi Angusowi, Krisowi Rammowi, a także Freydis Hjorvarsdottir (poprzednio CSP, obecnie Ministry of Primary Industries New Zealand) za wsparcie pracy. Podziękowania dla Helen Neil (NIWA) i Dana Sinclaira (VUW), za wsparcie w dostarczeniu ich wstępnych danych. Dziękujemy pracownikom NIWA: Sadie Mills i Dianie Macpherson za ich wkład w dobór próbek, Owenowi Andersonowi za przygotowanie Ryciny 1, Peterowi Hornowi za zapewnienie dokładnego przeglądu wewnętrznego oraz Rosemary Hurst za pomocne komentarze i porady dotyczące manuskryptu. Dziękujemy Jeremy Horowitzowi (James Cook University, Australia) za identyfikację próbek, oraz Dennisowi Opresko (Emeritus Smithsonian Institute) za komentarze dotyczące rewizji taksonomii koralowców czarnych. Finally, we thank the three reviewers, this document greatly benefitted from their detailed and constructive assessment.
Adkins, J. F., Griffin, S., Kashgarian, M., Cheng, H., Druffel, E. R. M., Boyle, E. A., et al. (2002). Radiocarbon dating of deep-sea corals. Radiocarbon 44, 567-580. doi: 10.1017/s0033822200031921
CrossRef Full Text | Google Scholar
Althaus, F., Williams, A., Schlacher, T. A., Kloser, R. J., Green, M. A., Barker, B. A., et al. (2009). Impacts of bottom trawling on deep-coral ecosystems of seamounts are long-lasting. Mar. Ecol. Prog. Ser. 397, 279-294. doi: 10.3354/meps08248
CrossRef Full Text | Google Scholar
Alves, E. Q., Macario, K., Ascough, P., and Bronk Ramsey, C. (2018). The worldwide marine radiocarbon reservoir effect: definitions, mechanisms, and prospects. Rev. Geophys. 56, 278-305. doi: 10.1002/2017RG000588
CrossRef Full Text | Google Scholar
Anderson, O., Tracey, D., Bostock, H., Williams, M., and Clark, M. (2014). Refined Habitat Suitability Modelling for Protected Coral Species in the New Zealand EEZ. NIWA Client Report przygotowany dla Department of Conservation. WLG2014-69No. 46. Auckland: NIWA.
Google Scholar
Andrews, A. H., Cailliet, G. M., Kerr, L. A., Coale, K. H., Lundstrom, C., and DeVogelaere, A. P. (2005). „Investigations of age and growth for three deep-sea corals from the Davidson Seamount off central California,” in Cold-Water Corals and Ecosystems, eds A. Freiwald and J. M. Roberts (Berlin: Springer-Verlag), 1021-1038. doi: 10.1007/3-540-27673-4_51
CrossRef Full Text | Google Scholar
Andrews, A. H., Cordes, E. E., Mahoney, M. M., Munk, K., Coale, K. H., Cailliet, G. M., et al. (2002). Age, growth and radiometric age validation of a deep-sea, habitat-forming gorgonian (Primnoa resedaeformis) from the Gulf of Alaska. Hydrobiologia 471, 101-110.
Google Scholar
Anthony, K. R. N., and Marshall, P. A. (2009). Coral Reefs and Climate Change. Marine Climate Change Impacts and Adaptation Report Card for Australia 2009. Gland: IUCN.
Google Scholar
Bronk Ramsey, C. (2001). Development of the radiocarbon calibration program OxCal. Radiocarbon 43, 355-363. doi: 10.1017/s0033822200038212
CrossRef Full Text | Google Scholar
Cairns, S., Gershwin, L., Brook, F., Pugh, P., Dawson, E., Ocanþa, O., et al. (2009). „Phylum cnidaria: corals, medusae, hydroids myxozo-ans,” in New Zealand Inventory of Biodiversity, Volume 1: Kingdom Animalia, ed. D. P. Gordon (New Zealand: Canterbury University Press), 59-101.
Google Scholar
Carreiro-Silva, M., Andrews, A. H., Braga-Henriques, A., de Matos, V., Porteiro, F. M., and Santos, R. S. (2013). Variability in growth rates of long-loved black coral Leiopathes sp. from the Azores. Mar. Ecol. Progr. Ser. 473, 189-199. doi: 10.3354/meps10052
CrossRef Full Text | Google Scholar
Chiswell, S. M., Bostock, H. C., Sutton, P. J. H., and Williams, M. J. M. (2015). Physical oceanography of the deep seas around New Zealand: a review, New Zealand. J. Mar. Freshw. Res. 49, 1-32. doi: 10.1080/00288330.2014.992918
CrossRef Full Text | Google Scholar
Clark, M., Bowden, D., Baird, S., and Stewart, R. (2010). Effects of fishing on the benthic biodiversity of seamounts of the „Graveyard” complex, northern Chatham Rise. New Zeal. Aqu. Environ. Biodivers. Rep. 46:40.
Google Scholar
Clark, M., and Rowden, A. (2009). Effects of deepwater trawling on the macro-invertebratebrate assemblages of seamounts on the Chatham Rise, New Zealand. Deep Sea Res. I 56, 1540-1554. doi: 10.1016/j.dsr.2009.04.015
CrossRef Full Text | Google Scholar
Clark, M., Tracey, D., Anderson, O., and Parker, S. (2014). Pilotażowa ocena ryzyka ekologicznego dla chronionych koralowców. Wellington: National Institute of Water and Atmospheric Research, 32.
Google Scholar
Clark, M. R., Bowden, D. A., Rowden, A. A., and Stewart, R. (2019). Little evidence of benthic community resilience to bottom trawling on seamounts after 15 years. Front. Mar. Sci. 6:63. doi: 10.3389/fmars.2019.00063
CrossRef Full Text | Google Scholar
Coleman, D. C. (1991). Carbon Isotope Techniques. Cambridge, MA: Academic Press.
Google Scholar
Consalvey, M., MacKay, K., and Tracey, D. (2006). Information review for protected deep-sea coral species in the New Zealand region. NIWA Client Report. WLG2006-85. 60. Auckland: NIWA.
Google Scholar
Druffel, E. R. M., Griffin, S., Witter, A., Nelson, E., Southon, J., Kashgarian, M., et al. (1995). Gerardia: bristlecone Pine of the deep-sea? Geochim. Cosmochim. Acta 59, 5031-5036. doi: 10.1016/0016-7037(95)00373-8
CrossRef Full Text | Google Scholar
Druffel, E. R. M., King, L. L., Belastock, R. A., and Buesseller, K. O. (1990). Growth rate of a deep-sea coral using 210Pb and other isotopes. Geochim. Cosmochim. Acta 54, 1493-1500.
Google Scholar
Fallon, S. J., Fifield, L. K., and Chappell, J. M. (2010). The next chapter in radiocarbon dating at the Australian National University: status report on the single stage AMS. Nuclear Instr. Methods Phys. Res. Sec. B 268, 898-901. doi: 10.1016/j.nimb.2009.10.059
CrossRef Full Text | Google Scholar
Freeman, D., and Cryer, M. (2019). „Current management measures and threats,” in The State of Knowledge of Deep-Sea Corals in the New Zealand Region, Chap. 9, eds D. M. Tracey and F. Hjorvarsdottir (Auckland: NIWA Science and Technology Series).
Google Scholar
Gammon, M. J., Tracey, D. M., Marriott, P. M., Cummings, V. J., and Davy, S. K. (2018). The physiological response of the deep-sea coral Solenosmilia variabilis to ocean acidification. PEERJ 6:e5236. doi: 10.7717/peerj.5236
PubMed Abstract | CrossRef Full Text | Google Scholar
Griffin, S., and Druffel, E. M. (1989). Sources of carbon to deep-sea corals. Radiocarbon 31, 533-543. doi: 10.1017/s0033822200012121
CrossRef Full Text | Google Scholar
Hitt, N. T., Sinclair, D. J., Fallon, S. J., Neil, H. L., Tracey, D. M., Komugabe-Dixson, A., et al. (2020). Growth and longevity of New Zealand black corals. Deep Sea Res. I 2020:103298. doi: 10.1016/j.dsr.2020.103298
CrossRef Full Text | Google Scholar
Komugabe, A. F., Fallon, S. J., Thresher, R. E., and Eggins, S. M. (2014). Modern Tasman Sea surface reservoir ages from deep-sea black corals. Deep-Sea Res. 99, 207-212. doi: 10.1016/j.dsr2.2013.05.024
CrossRef Full Text | Google Scholar
Komugabe-Dixson, A. F., Fallon, S. J., Eggins, S. M., and Thresher, R. E. (2016). Radiocarbon evidence for mid-late Holocene changes in southwest Pacific Ocean circulation. Paleoceanography 31, 971-985. doi: 10.1002/2016PA002929
CrossRef Full Text | Google Scholar
Love, M. S., Yoklavich, M. M., Black, B. A., and Andrews, A. H. (2007). Age of black coral (Antipathes dendrochristos) colonies, with notes on associated invertebratebrate species. Bull. Mar. Sci. 80, 391-400.
Google Scholar
Matsumoto, K. (2007). Radiocarbon-based circulation age of the world oceans. J. Geophys. Res. 112:C09004. doi: 10.1029/2007JC004095
CrossRef Full Text | Google Scholar
Mortensen, P. B., and Buhl-Mortensen, L. (2005). Morphology and growth of the deep-water gorgonians Primnoa resedaeformis and Paragorgia arborea. Mar. Biol. 147, 775-788. doi: 10.1007/s00227-005-1604-y
CrossRef Full Text | Google Scholar
Noé, S. U., Lembke-Jene, L., and Chr Dullo, W. (2008). Varying growth rates in bamboo corals: sclerochronology and radiocarbon dating of a mid-Holocene deep-water gorgonian skeleton (Keratosis sp.: Octocorallia) z Chatham Rise (Nowa Zelandia). Facies 54, 151-166. doi: 10.1007/s10347-007-0129-x
CrossRef Full Text | Google Scholar
Pinkerton, M. H. (2016). Ocean Colour Satellite Observations of Phytoplankton in the New Zealand EEZ, 1997-2016. Ministerstwo Środowiska. Wellington: NIWA.
Google Scholar
Prouty, N. G., Fisher, C. R., Demopoulous, A. W. J., and Druffel, E. R. M. (2016). Growth rates and ages of dee-sea corals impacted by the Deepwater Horizon oil spill. Deep-Sea Res. II 129, 196-212. doi: 10.1016/j.dsr2.2014.10.021
CrossRef Full Text | Google Scholar
Prouty, N. G., Roark, E. B., Buster, N. A., and Ross, S. W. (2011). Growth rate and age distribution of deep-sea black corals in the Gulf of Mexico. Mar. Ecol. Prog. Ser. 423, 101-115. doi: 10.3354/meps08953
CrossRef Full Text | Google Scholar
Reimer, P. J., Bard, E., Bayliss, A., Beck, J. W., Blackwell, P. G., Bronk Ramsey, C., et al. (2013). IntCal13 and Marine13 radiocarbon age calibration curves 0-50,000 years cal BP. Radiocarbon 55, 1869-1887. doi: 10.2458/azu_js_rc.55.16947
PubMed Abstract | CrossRef Full Text | Google Scholar
Reimer, P. J., and Reimer, R. W. (2001). A marine reservoir correction database and on-line interface. Radiocarbon 43, 461-463. doi: 10.1017/s0033822200038339
CrossRef Full Text | Google Scholar
Roark, E. B., Guilderson, T. P., Dunbar, R. B., Fallon, S. J., and Mucclarone, D. A. (2009). Extreme longevity in proteinaceous deep-sea corals. Proc. Natl. Acad. Sci. U.S.A. 106, 5204-5208. doi: 10.1073/pnas.0810875106
PubMed Abstract | CrossRef Full Text | Google Scholar
Roark, E. B., Guilderson, T. P., Dunbar, R. B., and Ingram, B. L. (2006). Radiocarbon based ages and growth rates: Hawaiian deepsea corals. Mar. Ecol. Progr. Ser. 327, 1-37.
Google Scholar
Sánchez, J., and Rowden, A. (2006). „Octocoral diversity on New Zealand Seamounts,” in Proceedings of the Presented at the 10th International Coral Reef Symposium, Okinawa.
Google Scholar
Sherwood, O. A., and Edinger, E. N. (2009). Ages and growth rates of some deep-sea gorgonian and antipatharian corals of Newfoundland and Labrador. Can. J. Fish. Aquatic Sci. 66, 142-152. doi: 10.1139/f08-195
CrossRef Full Text | Google Scholar
Sherwood, O. A., Scott, D. B., Risk, M. J., and Guilderson, T. P. (2005). Radiocarbon dowód dla rocznych pierścieni wzrostu w głębinowych oktokoral Primnoa resedaeformis. Mar. Ecol. Prog. Ser. 301, 129-134. doi: 10.3354/meps301129
CrossRef Full Text | Google Scholar
Sikes, E., Burgess, S., Grandpre, R., and Guilderson, T. (2008). Assessing modern deep-water ages in the New Zealand region using deep-water corals. Deep Sea Res. I 55, 38-49. doi: 10.1016/j.dsr.2007.10.004
CrossRef Full Text | Google Scholar
Stuiver, M., and Polach, H. A. (1977). Discussion reporting of 14C data. Radiocarbon 19, 355-363. doi: 10.1017/S0033822200003672
CrossRef Full Text | Google Scholar
Thresher, R., Rintoul, S. R., Koslow, J. A., Weidman, C., Adkins, J., and Proctor, C. (2004). Oceanic evidence of climate change in southern Australia over the last three centuries. Geophys. Res. Lett. 31:l07212.
Google Scholar
Thresher, R. E., MacRae, C. M., Wilson, N. C., and Gurney, R. (2007). Environmental effects on the skeletal composition of deep-water gorgonians (Keratoisis spp.; Isididae). Bull. Mar. Sci. 8, 409-422.
Google Scholar
Tracey, D., Bostock, H., and Shaffer, M. (2018). Ageing Methods for Protected Deep-Sea Corals: A Review and Recommendation for an Ageing Study. DOC Contract 4527 GMC – Age and Growth of coral (POP2017-07). NIWA Client Report WN2018035. Auckland: NIWA, 40.
Google Scholar
Tracey, D., Neil, H., Gordon, D., and O’Shea, S. (2003). Chronicles of the deep: ageing deep-sea corals in New Zealand waters. NIWA Water Atmosph. 11, 22-24.
Google Scholar
Tracey, D. M., and Hjorvarsdottir, F. (eds) (2019). „The state of knowledge of deep-sea corals in the new zealand region,” in NIWA Science and Technology Series Number 84, (Auckland: NIWA),140.
Google Scholar
Tracey, D. M., Neil, H., Marriott, P., Andrews, A. H., Cailliet, G. M., and Sanchez, J. A. (2007). Age and growth of two genera of deep-sea bamboo corals (family Isididae) in New Zealand waters. Bull. Mar. Sci. 81, 393-408.
Google Scholar
Williams, A., Schlacher, T. A., Rowden, A. A., Althaus, F., Clark, M. R., Bowden, D. A., et al. (2010). Seamount megabenthic assemblages fail to recover from trawling impacts. Mar. Ecol. 31, 183-199. doi: 10.1111/j.1439-0485.2010.00385.x
CrossRef Full Text | Google Scholar
Williams, B., and Grottoli, A. G. (2010). Stable nitrogen and carbon isotope (δ15N and δ13C) variability in shallow tropical Pacific soft coral and black coral taxa and implications for paleoceanographic reconstructions. Geochim. Cosmochim. Acta 74, 5280-5288. doi: 10.1016/j.gca.2010.06.026
CrossRef Full Text | Google Scholar
Williams, B., Risk, M. J., Ross, S. W., and Sulak, K. J. (2007). Stable isotope data from deep-water antipatharians: 400-year records from the southeastern coast of the United States of America. Bull. Mar. Sci. 81, 437-447.
Google Scholar
.
Leave a Reply