Frontiers in Marine Science
Introduction
Diepzeekoralen zijn een zeer diverse groep mariene organismen, en de regio Nieuw-Zeeland is een hotspot voor diepzeekoralen (Sánchez en Rowden, 2006; Cairns et al., 2009). Verschillende studies hebben de effecten van bodemtrawlvisserij op diepzee macro-invertebraten assemblages (inclusief koralen) in het zuidelijke deel van de Stille Oceaan onderzocht (Althaus et al., 2009; Clark and Rowden, 2009; Clark et al., 2010, 2019; Williams et al., 2010; Anderson et al., 2014). Deze hebben aangetoond dat de visserij met bodemtrawls aanzienlijke gevolgen heeft voor langzaam groeiende en kwetsbare diepzeekoraalkolonies en -gemeenschappen, die zich niet of nauwelijks kunnen herstellen. Milieueffecten die worden voorspeld als gevolg van klimaatverandering, zoals de opwarming van de oceanen en de verzuring van de oceanen, zullen naar verwachting ook een bedreiging vormen voor de gezondheid van diepzeekoralen, waarbij sommige habitats te warm worden voor hen om te gedijen in Anthony en Marshall (2009), of een verlaagde pH-waarde en carbonaationen zullen vertonen, wat waarschijnlijk gevolgen zal hebben voor diepzeekoralen met een op carbonaat gebaseerd skelet (Gammon et al., 2018). Om toekomstige risicobeoordelingen en het beheer van mariene hulpbronnen beter te informeren, is inzicht in de leeftijd en groei van diepzeekoralen essentieel voor het bepalen van de regeneratietijden van koralen en het herstelvermogen na antropogene verstoringen of andere natuurlijke schade.
Er is in de regio Nieuw-Zeeland veel werk verricht op het gebied van de taxonomie van diepzeekoralen, en de verspreiding van verschillende soorten en hun milieuvoorkeuren, evenals het belang van het diepzeekoraal voor de benthische ecosystemen. Een overzicht van ons begrip en onze kennis van diepzeekoralen in de regio Nieuw-Zeeland is onlangs gepubliceerd (Tracey en Hjorvarsdottir, 2019). In dit overzicht werden ook belangrijke hiaten in ons begrip van diepzeekoralen geschetst, waaronder beperkte leeftijds- en groeigegevens voor veel genera en soorten. In deze studie beoordelen we de leeftijd en groeisnelheid van het beschermde diepzee zwarte koraal Antipatharian Bathypathes patula. De leeftijd en de groeisnelheid van dit koraal zijn in deze regio nog niet eerder bestudeerd. De selectie van deze soort was gebaseerd op de prioriteitslijst van de risicobeoordeling (Clark et al., 2014) en de literatuurstudie (Tracey et al., 2018), gekoppeld aan de beschikbaarheid van monsters (locatie en totale aantallen), en ander aanvullend zwart koraalonderzoek dat in Nieuw-Zeeland wordt uitgevoerd (Hitt et al., 2020).
Tracey et al. (2018) schetsten de huidige methoden die worden gebruikt om diepzeekoraalsoorten te verouderen. De voor- en nadelen van elke methode werden besproken. De belangrijkste methoden die met succes zijn toegepast om leeftijd en groei van diepzeekoralen te meten, zijn (1) directe observatie, zoals in situ metingen of in-aquaria experimenten van lineaire groei of oppervlakte-uitbreidingen; toevoegingssnelheid van poliepen; schatting van calcificatiesnelheden (bijv, met behulp van de buoyant weight techniek), (2) opsomming van skeletale groeizones of banden (sclerochronologie), en (3) radiometrische analyses.
Directe waarnemingen en in situ of in-aquaria metingen zijn zeer moeilijk uit te voeren voor diepzeekoralen, omdat het moeilijk is om levende levensvatbare specimens te verkrijgen en in leven te houden omdat ze veel tijd nodig hebben om te groeien (Gammon et al., 2018; Tracey et al., 2018). Het opsommen van groeizones, Sclerochronologie, is ideaal voor diepzeekoraalsoorten die een groot contrast hebben tussen de zone-achtige groeipatronen die in skeletdoorsneden worden waargenomen. Het is echter essentieel dat een onafhankelijke validatiemethode wordt gebruikt om de chronologische geldigheid van de tellingen van de waargenomen groeizones te testen (Andrews et al., 2002). De methode om zichtbare groeizones in de skeletstructuur te verkrijgen is met succes toegepast om leeftijds- en groeisnelheidgegevens van gorgonische octokoralen te bepalen, waaronder 400 jaar oude bamboekoralen Keratoisis sp, 100 jaar oude zeekoralen Primnoa spp., en 60 jaar oude bamboekoralen Lepidisis sp. (Thresher et al., 2004, 2007; Mortensen and Buhl-Mortensen, 2005; Sherwood et al., 2005; Tracey et al., 2007; Sherwood and Edinger, 2009). Zwarte koralen zijn ook met succes verouderd aan de hand van groeizones of tellingen, met leeftijden van 150 jaar tot 480 jaar voor respectievelijk Antipathes dendrochristos en Leiopathes glaberrima (Love et al., 2007; Williams et al., 2007). In een studie over het zwarte koraal Stauropathes arctica werden zones van 55-58 jaar geteld (Sherwood en Edinger, 2009), waarbij de auteurs opmerkten dat de datering met behulp van radiokoolstof deze leeftijden beperkte tot 55 en 82 jaar. Groeisnelheden van zwarte koralen via de telling van groeizones onthullen een lage radiale groei van 8 tot 140 μm/jaar (Love et al., 2007; Prouty et al., 2011).
Er zijn potentiële beperkingen aan de telling van groeizone methode (zie samenvatting in Tracey et al., 2018). Veel koralen vertonen geen duidelijke regelmatige zonestructuur in hun hele skelet, of de duidelijke zonestructuur kan alleen duidelijk zijn in delen van het skelet. Zelfs wanneer aanwezig kan de zonestructuur variabel zijn of verschillende interpretatiemogelijkheden vertonen. Zo waren de basale en tiptellingen van bamboeoctokoraalsoorten Keratoisis sp. en Lepidisis sp. hoger dan de met de radiometrische lood 210 (210Pb) methode geschatte leeftijden (Tracey et al., 2007). Er werd verondersteld dat de zones die met een lichtmicroscoop werden waargenomen een tweejaarlijkse periodiciteit hebben en dat de met SEM waargenomen zones op het knooppunt een milieugebeurtenis kunnen vertegenwoordigen, zoals een maanperiodiciteit. Maancycli kunnen samengaan met sterke getijdengebeurtenissen, die de beschikbaarheid van voedsel voor sessiele filtervoedende organismen kunnen vergroten, wat kan leiden tot impulsen van somatische groei. Gorgonische bubblegum koralen (b.v. Paragorgia spp.) en rode edelsteenkoralen (b.v. Corallium sp.) zijn niet succesvol verouderd met behulp van zonetellingen vanwege de inherent complexe of slecht gedefinieerde bandpatronen (Griffin en Druffel, 1989; Andrews et al, 2005).
De meest gebruikte radiometrische methode bij diepzeekoralen is datering met het radiokoolstof (14C) isotoop (Druffel et al., 1995; Sherwood et al., 2005; Tracey et al., 2018). Deze methode berust op het feit dat een zeer kleine hoeveelheid natuurlijke koolstof in de atmosfeer radioactief is in de vorm van 14C en wordt opgenomen in het koraalskelet wanneer het zijn calciumcarbonaat of eiwit en chitine vormt (Adkins et al., 2002; Tracey et al., 2003; Consalvey et al., 2006). Omdat de halfwaardetijd van radiokoolstof bekend is als 5.730 jaar, kan deze methode, zij het duur, gebruikt worden om specimens van minder dan 50.000 jaar geleden betrouwbaar te verouderen; daarna wordt de activiteit van 14C te laag om te detecteren (Coleman, 1991). Tijdens de jaren 1950-60 was er een snelle toename van atmosferische 14C ten gevolge van het testen van kernwapens; deze “bomkoolstof” kan ook worden gebruikt als een instrument om de ouderdom te ijken, door een referentiepunt te verschaffen voor recentere monsters (Coleman, 1991; Druffel et al., 1995; Tracey et al., 2003; Roark et al, 2009; Sherwood and Edinger, 2009).
Radiokoolstofdatering is met succes gebruikt op verschillende koraalsoorten, waaronder steenkoralen, zwarte koralen en gorgonische octokoralen (Druffel et al., 1990; Roark et al., 2006; Carreiro-Silva et al., 2013; Prouty et al., 2016; Hitt et al., 2020). Er zijn echter ook beperkingen aan deze methode. Als gevolg van de oceaancirculatie is de radiokoolstofleeftijd van het water in de oceanen aanzienlijk ouder dan die in de atmosfeer, de zogenaamde “mariene reservoirleeftijd” (Tracey et al., 2003; Consalvey et al., 2006; Roark et al., 2006; Noé et al., 2008). Fouten in de reservoirleeftijd kunnen aanzienlijke gevolgen hebben voor de leeftijdsschattingen van koralen. Het corrigeren voor reservoirleeftijden vereist kennis van de omringende wateren en veronderstelt constante verhoudingen van radiokoolstof in het water; deze niveaus kunnen echter worden gewijzigd door vermenging binnen de oceaan, gelokaliseerde upwelling-evenementen en productiviteitsbloei, die allemaal de koolstofcyclustransformaties en koraalskeletsamenstelling van 14C kunnen veranderen (Tracey et al, 2018).
Op basis van de Tracey et al. (2018) review kozen we ervoor om twee verouderingsmethoden voor B. patula te gebruiken; tellingen van skeletgroeibanden en de radiometrische methode van radiokoolstofdatering (14C). De toepassing van deze methoden maakt vergelijkingen mogelijk met andere eerdere onderzoeken naar veroudering van koralen, zowel in Nieuw-Zeeland als wereldwijd. Monsterselectie, microfrezen van materiaal en sectioneringsmethoden, en de interpretatie van de zonetellingen en radiokoolstofdatering worden gepresenteerd. Leeftijds- en groeikarakteristieken van het zwarte koraal B. patula en één kolonie Bathypathes cf. conferta (waarvan aanvankelijk werd aangenomen dat het B. patula was) worden gegeven.
Materialen en methoden
Selectie van monsters
De monsters die in deze studie werden gebruikt, waren afkomstig van geconserveerde specimens die in het Nieuw-Zeelandse National Institute of Water and Atmospheric Research (NIWA), National Invertebrate Collection (NIC), zijn ondergebracht.
De monsters werden geselecteerd uit twee regio’s; de Bay of Plenty (ten noordoosten van het Noordereiland van Nieuw-Zeeland) en Chatham Rise (ten oosten van Nieuw-Zeeland), om de resultaten te vergelijken en de variabiliteit in groeisnelheid tussen verschillende gebieden te beoordelen, en ook met andere diepzee zwarte koraalsoorten, met name Antipathella, Leiopathes, en Antipathes, uit dezelfde regio’s (Hitt et al., 2020).
Oceanografische stromingen beïnvloeden diepzeekoralen door te zorgen voor een gestage stroom van voedsel en de verspreiding van larven. Bijgevolg beïnvloeden deze stromingen ook de beschikbaarheid van prooi en de verspreiding van diepzeekoralen. Terwijl diepzeekoralen het meest worden beïnvloed door de stromingen van het bodemwater die over hen heen stromen en de verspreiding van sedimenten beïnvloeden, worden zij ook beïnvloed door de stromingen van het oppervlaktewater die van invloed zijn op de primaire productiviteit en dus op de voedselvoorraad die naar het diepzeekoraal op de zeebodem neerdaalt. De landmassa van Nieuw-Zeeland bevindt zich op het kruispunt van warme, voedselarme, subtropische oppervlaktewateren die vanuit het noorden binnenstromen, en koele, voedselrijke, subantarctische oppervlaktewateren die vanuit het zuiden binnenstromen. Het huidige inzicht in de watermassa’s en oceaanstromingen in de regio aan de oppervlakte, thermocline, tussenliggende wateren, en diepe wateren, werd onlangs herzien door Chiswell et al. (2015). De oceanografische setting voor de koralen uit deze studie is als volgt.
De Bay of Plenty wordt omspoeld door warm, voedselarm, subtropisch oppervlaktewater dat afkomstig is van de East Australian Current en vanuit het westen binnenstroomt en de East Auckland Current vormt. De Chatham Rise is het ontmoetingspunt van het warme, subtropische water met een laag gehalte aan voedingsstoffen (STW) en het koele, subantarctische water met een hoog gehalte aan voedingsstoffen (SAW), dat afkomstig is van de Zuidelijke Oceaan en het Subtropische Front (STF) vormt. De vermenging van STW- en SAW-watermassa’s bij het STF leidt ertoe dat de Chatham Rise een zone is met een zeer hoge primaire productiviteit die een belangrijke voedselbron en dus koolstofvoorziening voor deze koralen vormt, chlorofyl a-concentraties variëren van 0,3 en 0,48 mg/m3 in de Chatham Rise en Bay of Plenty, vergeleken met 0,03-0,3 mg/m3 in de centrale Stille Oceaan (Pinkerton, 2016). Op een diepte van 500-1500 m stroomt het koele (3,5-10°C), lage zoutgehalte (34,4-34,6) en zuurstofrijke (200-250 μmol/kg) en nutriëntenrijke (nitraat 15-36 μmol/kg) Antarctische tussenwater (AAIW) vanuit het noorden en zuiden naar deze regio (Chiswell et al, 2015).
Koralen van de Chatham Rise werden gekozen omdat er moderne 14C-reservoirleeftijdsgegevens van dit gebied zijn (Sikes et al., 2008). Kennis van de plaatselijke 14C-reservoirleeftijd maakt een nauwkeuriger schatting van de radiokoolstofleeftijd van koralen mogelijk.
De monsters werden geselecteerd uit bestaande specimens die zijn verzameld door Fisheries New Zealand (FNZ), Ministry for Primary Industries (MPI) Observers en NIWA-onderzoekers en worden bewaard in het (NIC). Een plot van de verspreidingsgegevens voor B. patula heeft geholpen bij de beslissing over de keuze van de kolonieplaats en het aantal monsters (figuur 1). Twee specimens uit de Bay of Plenty (NIWA24190 en NIWA85940) werden geselecteerd voor radiokoolstofdatering en dunne doorsneden voor zonetellingen. De rest van de specimens werd geselecteerd uit de Chatham Rise.
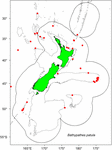
Figuur 1. Locaties waar Bathypathes patula is bemonsterd in de regio Nieuw-Zeeland. Rode stippen geven de locaties aan waar individuele koraalmonsters zijn genomen van B. patula-specimens die worden bewaard in de National Invertebrate Collection of New Zealand (NIC). Grijze lijnen geven de 200 m waterdieptecontour aan. Zwarte lijn geeft de omvang van de Nieuw-Zeelandse Exclusieve Economische Zone (EEZ) aan.
In het najaar van 2018 heeft een bezoekende specialist in de taxonomie van zwarte koralen (Jeremy Horowitz, James Cook University) de identificaties van verschillende monsters in het NIC herzien. Tijdens dit proces kreeg een van de zwarte koraalkolonies die voor dit leeftijdsschattingsonderzoek was geselecteerd en waarvan werd aangenomen dat het Bathypathes patula was, een herziene identificatie. Het zwarte koraal NIWA24190 is nu geïdentificeerd als Bathypathes cf. conferta. Het monster is in dit artikel aangeduid als Bathypathes cf. conferta (figuur 2), de meest waarschijnlijke correcte identificatie. Verdere taxonomische beoordeling van dit specimen is gaande (Opresko pers comm). De verspreidingskaart (Figuur 1) kan bijkomende exemplaren van B. cf. conferta bevatten.

Figuur 2. Bathypathes cf. conferta. NIWA24190. Exemplaar dat laat zien waar monsters zijn genomen voor analyse. (A) is de groeitop, (B) de basale doorsnede. Exemplaar uit Bay of Plenty, Nieuw-Zeeland. De schaalbalk is een liniaal van 35 cm.
Voorbereiding van het materiaal
Eenmaal geschikte monsters waren geïdentificeerd, werden kleine (minder dan 10 mm secties) uit de groeitoppen en het basale gedeelte van de hoofdsteel van de koralen gesneden. Aangezien veel van de monsters tijdens het verzamelen waren gebroken, werden extra secties genomen langs de hoofdstengel, zodat betrouwbare schattingen van de lineaire groeisnelheid konden worden gegenereerd (figuren 2-5). De monsters van de groeitoppen werden vervolgens in twee fragmenten gesplitst, één voor radiokoolstofdatering, het andere voor de preparatie van dunne doorsneden om zonetellingen te verkrijgen.

Figuur 3. Bathypathes patula NIWA85940. Exemplaar waarop te zien is waar monsters voor analyse zijn genomen. (A) is de groeitop, (C) de basale doorsnede, en (B) is een tussenliggende monsterplaats aangezien het specimen bij het verzamelen gebroken was en er boven deze plaats onbekende lengtes van de hoofdtakstengel kunnen ontbreken. Exemplaar uit Bay of Plenty, Nieuw-Zeeland. Schaalbalk is een liniaal van 35 cm.

Figuur 4. Bathypathes patula NIWA49468. Exemplaar waarop te zien is waar monsters voor analyse zijn genomen. (A) is de groeitop, (D) de basale doorsnede, en (B,C) zijn tussenliggende monsterplaatsen aangezien het specimen bij het verzamelen gebroken was en een onbekende lengte van de hoofdtakstengel tussen deze twee plaatsen kan ontbreken. Exemplaar uit Chatham Rise, Nieuw-Zeeland. Schaalbalk is een liniaal van 35 cm.

Figuur 5. Bathypathes patula NIWA47911. Exemplaar dat toont waar monsters werden genomen voor analyse. (A) is de groeitop, (F) de basale doorsnede en, (B-E) zijn tussenliggende monsterplaatsen genomen om te zoeken naar het radiokoolstofbommensignaal. Exemplaar van Chatham Rise, Nieuw-Zeeland. Schaalbalk is een 35 cm liniaal.
Micro-Milling of Material
Vanaf eerder werk was opgemerkt dat zwarte koraalsecties vatbaar zijn voor het omhoog trekken van hars door hun poreuze matrix via capillaire werking en dus konden de secties in deze studie niet met hars op een grondplaat worden vastgelijmd om te frezen. In plaats daarvan werd een houder vervaardigd om de doorsnede tijdens het frezen vast te houden, waardoor het risico van harsbesmetting van de radiokoolstofmonsters werd geëlimineerd.
Gepoederde radiokoolstofmonsters werden uit de rand en de kern van basale, middelste en topdoorsneden geëxtraheerd met behulp van een New WaveTM micromolen met een 0,5 mm Brassler H2.11.006 freesbraam van het NIWA. De randmonsters bestonden uit materiaal van de buitenste 250 micron van de doorsnede, de kernmonsters werden verkregen uit materiaal binnen een straal van 750 micron van het primordium. We hebben geprobeerd om per monster ten minste 1,5 mg materiaal te extraheren. De monsters werden vervolgens gewogen en geanalyseerd op radiokoolstof in het Radiocarbon Laboratorium van de Australian National University (ANU).
Dunne Sectie Methode en Protocollen voor Zone Telling
De doorsneden van de stam van het koraal werden ingebed in doorzichtig tweedelig epoxyhars en bij het NIWA doorgezaagd met een diamanten schaafzaag. De secties werden aan één zijde gepolijst en met de gepolijste zijde naar beneden op een glazen microscoopglaasje gemonteerd, vervolgens geslepen en gepolijst tot ze een optimale dikte hadden voor het bekijken van de groeizone-structuur met behulp van doorvallend licht. De optimale dikte voor coupes van deze soort was ongeveer 250-300 μm en volgt de standaardprocedure voor het prepareren van dunne coupes (Andrews et al., 2002; Mortensen and Buhl-Mortensen, 2005; Tracey et al., 2007). Het tellen van de periodieke groeibanden of incrementen, waarvan wordt aangenomen dat ze jaarlijks worden gevormd, vond plaats na het dunsnede prepareren bij NIWA.
Om zonetellingen te genereren, werden dunsnede preparaten van de hoofdtaksecties bekeken onder een samengestelde microscoop met doorvallend licht. Als hulpmiddel bij de zone-interpretatie werden de coupes ook bekeken met ultraviolet licht volgens de methodologie van Sherwood en Edinger (2009). Hier werden de zones geteld met behulp van doorvallend helderveldlicht en was ultraviolette verlichting een nuttig hulpmiddel om de waargenomen zonestructuur verder te helpen definiëren.
Radiokoolstofdateringsmethode
Vóór de analyses werden de monsters gereinigd (bv. met zuur uitgeloogd) van zwarte korsten en endolithische activiteit om alle mogelijke jongere verontreinigende 14C te verwijderen die de resultaten kunnen veranderen (Adkins et al., 2002). Poedervormige koraalmonsters werden behandeld met <1 ml 0,1 M HCL. Het zuur werd vervolgens weggepipetteerd en de monsters werden vervolgens driemaal gewassen met milliQ water. Het monster werd vervolgens bevroren met droog ijs en gelyofiliseerd. Ongeveer 1 mg schoongemaakt koraalmonster werd afgewogen in een 6 mm OD kwartsbuisje, waaraan ∼60 mg CuO en een zilvercupje werd toegevoegd. De buisjes werden geëvacueerd tot <3e-3 torr (<0,4 Pa) en verzegeld met een toorts. De buizen werden vervolgens gedurende 6 uur bij 900°C gebakken om CO2 te genereren. Het resulterende CO2 werd gezuiverd en omgezet in grafiet in aanwezigheid van waterstof met Fe als katalysator. Het grafiet werd vervolgens gemeten op de ééntraps versneller massaspectrometer van de ANU (Fallon et al., 2010). Alle gegevens zijn gecorrigeerd met behulp van on-line AMS δ13C, genormaliseerd naar oxaalzuur I en van de achtergrond afgetrokken met behulp van 14C-vrije steenkool die op dezelfde manier als de koraalmonsters is behandeld. De gegevens worden gepresenteerd volgens de aanbevelingen van Stuiver en Polach (1977).
Resultaten
Dunne secties voor zonetellingen
De zone-structuurpatronen die op de skeletsecties voor deze soorten zijn waargenomen, zijn zeer complex en kunnen op vele manieren worden geïnterpreteerd. Voor de radiale groeitellingen werden de geprepareerde doorsneden zorgvuldig geobserveerd en de as die het best een duidelijke zone-structuur vertoonde van het primordium tot de buitenrand werd vervolgens gebruikt om de zonetellingen te verkrijgen. De radiale groei van de takken in B. patula was redelijk regelmatig, er was geen duidelijke bias in de zone-afzetting die dikkere zones vormde aan een bepaalde kant van de tak, wat eerder is waargenomen bij andere zwarte koraalsoorten. De stralen gebruikt voor de groeisnelheid berekeningen was een gemiddelde van de minimale en maximale stralen waargenomen in de geprepareerde secties, wat betekent dat de gerapporteerde radiale groeisnelheden zal een gemiddelde radiale groeisnelheid voor het koraal.
Op het koraal skelet is er een eerste grove band patroon grotendeels gedefinieerd door afwisselend donkerder en lichtere zones (figuur 6) wanneer waargenomen onder lager vermogen (10-20×). Bij nadere beschouwing onder hoger vermogen (100-200×) is er ook een redelijk regelmatig fijnbandig patroon zichtbaar (figuur 7). Hier hebben we twee afleesprotocollen toegepast – waarbij zowel de grof gevormde brede zones als de dunne fijn gevormde zones zijn geteld (tabel 1). De hier gepresenteerde zonetellingen zijn allemaal verricht door één enkele lezer (Marriott). Tijdens de ontwikkeling van de protocollen voor zone-interpretatie werden vele metingen verricht met toepassing van de verschillende iteraties van de protocollen die werden ontwikkeld. De zonetellingen van de twee hier gepresenteerde protocollen werden elk afgeleid van meerdere tellingen over de doorsnede.

Figuur 6. Basale doorsnede van koraal NIWA47911 met de grof gekleurde bandinterpretatie van de waargenomen zone-structuur. Het specimen vertoont 54 zones, gemarkeerd met rode stippen. De gestippelde rechthoek binnenin toont het centrale gedeelte van de basale doorsnede die is afgebeeld in Figuur 7.

Figuur 7. Centraal gedeelte van de basale doorsnede van koraal NIWA47911 met het protocol voor de fijnschalige interpretatie van de waargenomen zonestructuur. Het specimen vertoont 42 zones, gemarkeerd met zwarte stippen in dit binnenste gedeelte van de doorsnede; de gehele doorsnede vertoonde 211 zones.

Tabel 1. Zone-tellingen van dunne doorsneden.
Voor het jongste deel van de bestudeerde specimens, de top, of nabij de top regio, varieerden de tellingen van 4 tot 7 (grove zone tellingen) en 10 tot 19 (fijne zone tellingen). In het middengebied van de hoofdtak varieerden de tellingen van 55 tot 60 (tellingen in de grove zone) en 370 tot 374 (tellingen in de fijne zone). Voor de basisregio van de kolonie varieerden de tellingen van 37 tot 193 (tellingen van de grove zone) en van 78 tot 481 (tellingen van de fijne zone). De complexiteit van de zoneringspatronen onderstreepte de noodzaak van validatie.
Radiokoolstofanalyse
De in tabel 2 vermelde gemalen monsters werden naar ANU gezonden voor radiokoolstofdatering. De resultaten van de 17 koraalmonsters zijn weergegeven in de radiokoolstofresultaten (Tabel 3). Drie monsters (monsters 47911-F2; 24190-B1; 85940-B1) zijn in het laboratorium mislukt; er was onvoldoende materiaal over om een duplicaat te proberen. De radiokoolstofresultaten van de resterende koralen werden gekalibreerd naar een kalenderleeftijd met behulp van OxCal4.3 (Bronk Ramsey, 2001), de Marine13-curve en een Nieuw-Zeelandse lokale deltaR (radiokoolstofreservoir-offset) van -18 ± 36 (Reimer en Reimer, 2001). De verdeling van de leeftijdsgrenzen voor koraalmonsters van vóór de bomaanslag is weergegeven in Tabel 3. Alle koraalspecimens die in deze studie werden gebruikt, waren levend bij de vangst, dus alle leeftijdsschattingen zijn de leeftijd bij de vangst en niet de levensduur van het specimen. De verwachte levensduur van deze soort zal hoger liggen dan de hier vermelde leeftijdsschattingen. Koraal 85940 was ongeveer 380 ± 50 jaar oud, koraal 49468 ongeveer 310 ± 70 jaar, en koraal 47911 ongeveer 120 ± 50 jaar (tabel 3). Alle monsters van de koraalbuitenrand en -toppen bevatten radiokoolstofwaarden F14C >1 (Tabel 3), behalve voor 85940-B2 en 85940-C2, wat erop wijst dat deze koralen nog leefden en oppervlaktewaterkoolstof opnamen op het ogenblik van de verzameling. Koralen 85940-B2 en 85940-C2 hadden radiokoolstofwaarden F14C <1 aan de buitenrand (Tabel 3), wat erop wijst dat een deel van het skelet niet actief materiaal toevoegde. Dit is eerder waargenomen waarbij het basale deel van de koraalkolonie stopt met groeien, het bovenste deel blijft echter doorgroeien (Komugabe-Dixson et al., 2016).
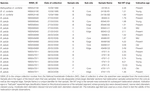
Tabel 2. Monsters geëxtraheerd voor radiokoolstofdatering.

Tabel 3. Resultaten van de radiokoolstofdatering.
Raming van de groeisnelheid
Raming van de groeisnelheid werd gebaseerd op de resultaten van de radiokoolstofdatering, aangezien tellingen van zones problematisch waren en niet gevalideerd bleken te zijn, zodat ze niet konden worden gebruikt om betrouwbare schattingen van de groeisnelheid te genereren (zie de paragraaf “Leeftijds- en groeischattingen voor Bathypathes patula” voor een verdere bespreking van de telling van zones). Groeisnelheden werden afgeleid uit de fysieke afstanden tussen de monsterpunten en de gekalibreerde koolstofleeftijden op de desbetreffende monsterplaats. Omdat sommige monsters tijdens de radiokoolstofanalyse mislukten, kon slechts een beperkte reeks schattingen van de groeisnelheid worden gegenereerd. Radiale groeisnelheden, de snelheid waarmee de kolonietak in de loop van de tijd radiaal dikker wordt, werden geschat door de gekalibreerde kalenderjaren van groei tussen de kern en de rand van de bemonsteringsplaats te delen door de gemiddelde straal van het taksegment. De radiale groei varieerde van 11,1 tot 35,7 μm/jaar. Lineaire groeisnelheden, de snelheid waarmee de kolonietak in de loop van de tijd in de lengterichting wordt verlengd, werden geschat door het verschil tussen de gekalibreerde kalenderleeftijden van de kernmonsters te delen door de lineaire afstand tussen twee monsterpunten. Twee lineaire groeisnelheden werden verkregen, 5,2 en 9,6 mm/jaar (Tabel 4).

Tabel 4. Bathypathes patula growth rate estimates.
Discussie
Bathypathes patula Age and Growth Estimates
De radiokoolstofresultaten van dit werk tonen aan dat B. patula een langlevende soort is, met leeftijden van meer dan 385 jaar. Uit de radiokoolstofleeftijdsgegevens werden langzame lineaire (5,2-9,6 mm/jaar) en radiale (11,1-35,7 μm/jaar) groeisnelheden verkregen.
Geen van de protocollen voor het tellen van zones correleerde betrouwbaar met de onafhankelijke schattingen van de radiokoolstofleeftijd. Het protocol voor het tellen van de fijne zones vertoonde de nauwste correlatie met de 14C-leeftijdsgegevens, wat enige algemene ondersteuning biedt voor de levensduur bij vergelijking van de zonetellingen met 14C-data. De leeftijdsschattingen tussen de radiokoolstofleeftijdsgegevens en de fijnschalige zonetellingen vertoonden routinematig een verschil van meer dan honderd jaar in taksecties met gekalibreerde radiokoolstofleeftijden van 112-385 jaar (Tabellen 1, 3). Verschillen in geschatte ouderdom van vergelijkbare monsters met behulp van de twee methoden waren negatief en positief; er was geen onder- of over-oudering met behulp van zonetellingen met een evenredige hoeveelheid (Tabel 1). Dit betekent dat de waargenomen zone-structuur geen jaarlijkse, noch een ander regelmatig chronologisch patroon van periodiciteit vertoont, en dat onze protocollen voor het tellen van zones niet kunnen worden gebruikt om op betrouwbare wijze leeftijdsschattingen voor B. patula te genereren.
De radiokoolstof-resultaten van de rand van specimen NIWA-85940 suggereren dat de basale sectie (C2) van de hoofdstengel 293 jaar voor de oogst ophield met het afzetten van skeletmateriaal op het buitenoppervlak, en dat de middensectie (B2) 88 jaar voor de oogst ophield met het afzetten van skeletmateriaal. Dit ondanks de waarneming dat deze delen van de vertakte stengel onmiddellijk grensden aan laterale pinnules met levende poliepen. Het optische aspect van deze secties gaf geen indicatie dat de afzetting van de zone aan hun buitenrand was gestopt. Het oppervlak van de koraaltak bleek nog relatief ongerept te zijn, het vertoonde geen tekenen van oppervlakkige erosie of verkleuring, zoals te verwachten zou zijn bij een koraaltak die niet langer nieuw skeletmateriaal aan het buitenoppervlak aangroeit. Dit suggereert dat als takstammen kunnen stoppen met het aangroeien van skeletlagen, terwijl de uiteinden van de tak blijven groeien, de protocollen van de zonetelmethode niet betrouwbaar kunnen worden gebruikt voor het genereren van leeftijdsschattingen van basale doorsneden voor deze koraalsoort.
In specimen NIWA47911 werden vijf monsters genomen van het groeiende uiteinde van de belangrijkste axiale tak naar de basis van het koraal. Deze monsters werden op een afstand van 25 mm van elkaar genomen. Dit was een poging om het verschijnen van bomradiokoolstof in het skelet van het koraal te lokaliseren. Dit zou betekenen dat de lineaire groeisnelheid nauwkeurig kon worden bepaald voor de recentere groeitoppen van de takken van de kolonie. Als de stijging van het bom-röntgencarbonaat nauwkeurig kon worden bepaald, zou dat ook een nauwkeurig referentiepunt opleveren voor de vaststelling van de 14C-correctie die moet worden toegepast op de monsters van vóór de bomaanslag voor deze kolonie en op andere kolonies uit dezelfde watermassa. Helaas bleken al deze monsters van na de bomaanslag te zijn, zodat dit proces niet kon worden voltooid. Toekomstige bemonstering verder terug in deze vertakking zou nuttig zijn om dit knooppunt en de hedendaagse lineaire groeisnelheid vast te stellen. In ons werk werd een lineaire groei gemeten van 5,2-9,6 mm/jaar, uitgaande van de snelste groeisnelheid zou je vanaf de groeitop 700 mm terug langs de hoofdtak moeten bemonsteren om er zeker van te zijn dat de toename van bomradiokoolstof in het skelet kan worden waargenomen.
Onderzoek van Love et al. (2007) aan de zwarte koraalsoort Antipathes dendrochristos meldde ook twee interpretaties van de zone-structuur, vergelijkbaar met de hier beschreven grove en fijne protocollen. Deze auteurs vonden echter dat de grover gekleurde bandstructuur correleerde met hun radiokoolstof- en 210Pb-resultaten die wijzen op een jaarlijkse bandering (Love et al., 2007).
De radiokoolstofmetingen van het huidige werk vertoonden F14C >1 in monsters van de groeiende takuiteinden en het hoofd-takoppervlak, wat erop wijst dat deze monsters moderne post-bom niveaus van radiokoolstof vertoonden. De aanwezigheid van post-bom koolstof onder de gemengde laag van de oceaan, geeft aan dat de B. patula zwarte koralen gebruik maken van recent geëxporteerde oppervlaktewater POC. Op deze diepten is POC de enige koolstofbron die met bom 14C is gemerkt. De opgeloste anorganische koolstof (DIC) van het omgevingswater waarin de koralen leven, heeft veel lagere 14C-waarden (Matsumoto, 2007; Alves et al., 2018). Het detecteren van bom 14C in koralen die groeien in wateren die geen bom 14C hebben ervaren (d.w.z. door oceaanventilatie) versterkt dat zwarte koralen hun koolstof ontlenen aan oppervlakte-afgeleide POC (Roark et al., 2009; Sherwood en Edinger, 2009; Williams en Grottoli, 2010; Prouty et al., 2011). Dit betekent ook dat we geen model hoeven te maken voor een diepwaterreservoireffect, zoals het geval is bij de steenkoralen, die hun skeletkoolstof ontlenen aan DIC in de tussenliggende wateren waarin ze baden.
Vergelijkingen met recente Nieuw-Zeelandse en wereldwijde leeftijdsgegevens van zwarte koralen
De uit radiokoolstof afgeleide leeftijdsschattingen en groeisnelheden van dit project zijn vergelijkbaar met de resultaten van een aantal andere leeftijds- en groeistudies die recent zijn ondernomen voor belangrijke diepzee zwarte koralen (Antipatharia) in de regio Nieuw-Zeeland (tabel 5). Deze leeftijds- en groeischattingen voor B. patula zijn vergelijkbaar met andere koraalgeslachten, zoals Leiopathes, Antipathes, en Antipathella kolonies, die tot 2600 jaar leven en groeien in de orde van 1-143 μm/jaar (Hitt et al., 2020). Onze resultaten ondersteunen verder dat Nieuw-Zeelandse zwarte koralen tot de langzaamst groeiende zeedieren in de regio behoren, waarbij sommige soorten een extreme levensduur vertonen.

Tabel 5. De resultaten van B. patula en andere zwarte koralen die in Nieuw-Zeeland zijn verouderd, laten geen duidelijke trends zien in groeisnelheid of levensduur met betrekking tot de watermassa’s en diepten waar deze koralen voorkomen. Monsters van de Chatham Rise en Bay of Plenty kwamen meestal van redelijk gelijke waterdieptes, tussen 750 en <1269 m. Het dieper gelegen Chatham Rise B. patula specimen kwam van ergens tussen 1120 en 1269 m (Tabellen 1, 5). De radiale groeisnelheid van onze Chatham Rise-monsters (NIWA-49468 en NIWA47911) was met 20-36 μm/jaar hoger dan die van het Bay of Plenty-exemplaar (NIWA-85940) met 11 μm/jaar. Aangezien slechts drie groeisnelheden met succes werden verkregen voor de B. patula monsters uit deze twee regio’s, kunnen geen veronderstellingen worden gemaakt over de regionale verschillen in groeisnelheid. Uit het spectrum van groeisnelheidsgegevens voor zwarte koralen van Chatham Rise blijkt dat een Antipathes sp. monster de snelste tot dusver waargenomen groeisnelheid had, 91-143 μm/jaar, en een van de traagste, 5-15 μm/jaar voor een Leiopathes secunda specimen. De zwarte koraalsoorten Bathypathes patula, Leiopathes secunda en Leiopathes sp. in de Bay of Plenty vertonen een gemiddelde tot lage radiale groeisnelheid, 5-40 μm/jaar. Daarentegen heeft het ondiepwater Fiordland zwart koraal Antipathella fiordensis een relatief hoge radiale groeisnelheid (50-57 μm/jr), maar het is onduidelijk of dit een functie is van de soort of van de milieuomstandigheden in het Fiord, of een mengeling van beide.
Dit onderzoek komt ook overeen met wereldwijde studies die hebben aangetoond dat zwarte koralen tot de langzaamst groeiende diepzeekoralen behoren. Sherwood en Edinger (2009) geven leeftijdsschattingen voor het zwarte koraal Stauropathes arctica tot 82 kalenderjaren. Prouty et al. (2011) gebruikten radiokoolstof om leeftijdsschattingen te genereren van levende bijvangstkolonies van Leiopathes sp. tot 2100 jaar. Carreiro-Silva et al. (2013) hebben een Azorenkolonie van Leiopathes sp. op 2320 jaar oud geschat. Een exemplaar van Leiopathes annosahas uit Hawaii vertoonde het potentieel om een levensduur van meer dan 4000 jaar te bereiken (Roark et al., 2009).
Deze nieuwe leeftijds- en groeicijfers voor het delicate diepzee zwarte koraal B. patula benadrukken dat dit een langlevende soort is, waarvan in deze studie is aangetoond dat ze leeftijden van meer dan 385 jaar bereikt, en langzame lineaire en radiale groeisnelheden vertoont. De resultaten wijzen op een waarschijnlijk geringe veerkracht en een langzame hersteltijd na verstoring/verwijdering. Dit onderzoek zal worden gebruikt om toekomstige risicobeoordelingen en beheerstrategieën in het Nieuw-Zeelandse kader voor het beheer van koralen te onderbouwen (Freeman en Cryer, 2019). De langzame groei en extreme levensduur benadrukt verder dat deze koralen zeer kwetsbaar zullen zijn voor onderzeese verstoringen, zoals visserij en mijnbouw, en er eeuwen over kunnen doen om te herstellen van antropogene invloeden (Hitt et al, 2020).
Conclusie
De leeftijd en groeisnelheid van diepzee zwart koraal B. patula monsters uit twee regio’s van Nieuw-Zeeland, Bay of Plenty en Chatham Rise werden bepaald met behulp van twee verschillende methoden; het tellen van groeiringen (sclerochronologie) en radiokoolstofdatering. Er werden twee verschillende protocollen gebruikt om de groeiringen te tellen, waarbij gebruik werd gemaakt van donkere en lichte groeilagen. Dit leverde uiteenlopende resultaten op, die ook niet in overeenstemming waren met de koolstofleeftijden van verschillende van deze koralen. De koolstofleeftijden van B. patula geven aan dat deze koraalsoort lang leeft, met leeftijden van meer dan 385 jaar, met langzame lineaire (5,2-9,6 mm/jaar) en radiale (11,1-35,7 μm/jaar) groeisnelheden. Deze resultaten zijn vergelijkbaar met de leeftijden en groeisnelheden van andere zwarte koraalsoorten uit dezelfde regio’s van Nieuw-Zeeland en wereldwijd. Leeftijds- en groeisnelheden van diepzeesoorten zijn van cruciaal belang om het potentiële risico van deze organismen voor verstoringen door de mens te bepalen. Deze resultaten zullen worden gebruikt om toekomstige risicobeoordelingen en beheerstrategieën in het Nieuw-Zeelandse kader voor het beheer van koralen te informeren.
Data Availability Statement
Alle datasets die voor deze studie zijn gegenereerd, zijn opgenomen in het artikel/aanvullend materiaal.
Author Contributions
PM, DT, HB, en NH hebben bijgedragen aan de conceptie en het ontwerp van de studie. PM heeft alle monstervoorbereidingen, de beeldanalyse van de coupes, de ontwikkeling van het protocol voor dunne coupes en het tellen van de zones, en het micromalen van de radiokoolstofmonsters uitgevoerd. NH analyseerde de radiokoolstofmonsters. SF maakte gecorrigeerde leeftijdsschattingen van de ruwe radiokoolstofgegevens en schreef delen van het manuscript. PM en DT schreven de eerste versie van het manuscript. Alle auteurs hebben bijgedragen aan de revisie van het manuscript.
Funding
Dit onderzoek werd gefinancierd uit jaar twee van het Conservation Services Programme (CSP), Department of Conservation (DOC) Project POP2017-07 Doelstelling: “Ontwikkeling van een methodologie om de leeftijd en groeikenmerken van belangrijke Nieuw-Zeelandse diepzee (koudwater) koraalsoorten met een hoog risico te bepalen.” Financiering van de publicatievergoeding voor dit tijdschrift werd verstrekt door NIWA, Nieuw-Zeeland.
Conflict of Interest
De auteurs verklaren dat het onderzoek werd uitgevoerd in afwezigheid van commerciële of financiële relaties die zouden kunnen worden opgevat als een potentieel belangenconflict.
Aankondigingen
Wij danken de Conservation Services Programme Group, Department of Conservation New Zealand, in het bijzonder Ian Angus, Kris Ramm, en ook Freydis Hjorvarsdottir (voorheen van CSP nu Ministry of Primary Industries New Zealand) voor hun steun aan het werk. Dank aan Helen Neil (NIWA) en Dan Sinclair (VUW), voor hun steun bij het verstrekken van hun voorlopige gegevens. Wij erkennen NIWA medewerkers Sadie Mills en Diana Macpherson voor hun inbreng in de steekproefselectie, Owen Anderson voor de productie van figuur 1, Peter Horn voor het verstrekken van een grondige interne review, en Rosemary Hurst voor nuttig commentaar en advies over het manuscript. Wij danken Jeremy Horowitz (James Cook University, Australië) voor de identificatie van de monsters, en Dennis Opresko (Emeritus Smithsonian Institute) voor commentaar op herzieningen van de taxonomie van zwart koraal. Tenslotte danken wij de drie reviewers, dit document heeft veel baat gehad bij hun gedetailleerde en constructieve beoordeling.
Adkins, J. F., Griffin, S., Kashgarian, M., Cheng, H., Druffel, E. R. M., Boyle, E. A., et al. (2002). Radiokoolstofdatering van diepzeekoralen. Radiocarbon 44, 567-580. doi: 10.1017/s0033822200031921
CrossRef Full Text | Google Scholar
Althaus, F., Williams, A., Schlacher, T. A., Kloser, R. J., Green, M. A., Barker, B. A., et al. (2009). Impacts of bottom trawling on deep-coral ecosystems of seamounts are long-lasting. Mar. Ecol. Prog. Ser. 397, 279-294. doi: 10.3354/meps08248
CrossRef Full Text | Google Scholar
Alves, E. Q., Macario, K., Ascough, P., and Bronk Ramsey, C. (2018). Het wereldwijde mariene radiokoolstofreservoireffect: definities, mechanismen, en vooruitzichten. Rev. Geophys. 56, 278-305. doi: 10.1002/2017RG000588
CrossRef Full Text | Google Scholar
Anderson, O., Tracey, D., Bostock, H., Williams, M., and Clark, M. (2014). Refined Habitat Suitability Modelling for Protected Coral Species in the New Zealand EEZ. NIWA Client Report opgesteld voor Department of Conservation. WLG2014-69No. 46. Auckland: NIWA.
Google Scholar
Andrews, A. H., Cailliet, G. M., Kerr, L. A., Coale, K. H., Lundstrom, C., and DeVogelaere, A. P. (2005). “Investigations of age and growth for three deep-sea corals from the Davidson Seamount off central California,” in Cold-Water Corals and Ecosystems, eds A. Freiwald and J. M. Roberts (Berlin: Springer-Verlag), 1021-1038. doi: 10.1007/3-540-27673-4_51
CrossRef Full Text | Google Scholar
Andrews, A. H., Cordes, E. E., Mahoney, M. M., Munk, K., Coale, K. H., Cailliet, G. M., et al. (2002). Age, growth and radiometric age validation of a deep-sea, habitat-forming gorgonian (Primnoa resedaeformis) from the Gulf of Alaska. Hydrobiologia 471, 101-110.
Google Scholar
Anthony, K. R. N., and Marshall, P. A. (2009). Koraalriffen en klimaatverandering. Rapport over de effecten van klimaatverandering op zee en aanpassing voor Australië 2009. Gland: IUCN.
Google Scholar
Bronk Ramsey, C. (2001). Ontwikkeling van het radiokoolstofkalibratieprogramma OxCal. Radiocarbon 43, 355-363. doi: 10.1017/s0033822200038212
CrossRef Full Text | Google Scholar
Cairns, S., Gershwin, L., Brook, F., Pugh, P., Dawson, E., Ocanþa, O., et al. (2009). “Phylum cnidaria: corals, medusae, hydroids myxozo-ans,” in New Zealand Inventory of Biodiversity, Volume 1: Kingdom Animalia, ed. D. P. Gordon (Nieuw-Zeeland: Canterbury University Press), 59-101.
Google Scholar
Carreiro-Silva, M., Andrews, A. H., Braga-Henriques, A., de Matos, V., Porteiro, F. M., and Santos, R. S. (2013). Variabiliteit in groeisnelheden van langlevend zwart koraal Leiopathes sp. van de Azoren. Mar. Ecol. Progr. Ser. 473, 189-199. doi: 10.3354/meps10052
CrossRef Full Text | Google Scholar
Chiswell, S. M., Bostock, H. C., Sutton, P. J. H., and Williams, M. J. M. (2015). Fysische oceanografie van de diepe zeeën rond Nieuw-Zeeland: een overzicht, New Zealand. J. Mar. Freshw. Res. 49, 1-32. doi: 10.1080/00288330.2014.992918
CrossRef Full Text | Google Scholar
Clark, M., Bowden, D., Baird, S., and Stewart, R. (2010). Effecten van visserij op de bentische biodiversiteit van onderzeese bergen van het “Graveyard” complex, noordelijke Chatham Rise. New Zeal. Aqu. Environ. Biodivers. Rep. 46:40.
Google Scholar
Clark, M., and Rowden, A. (2009). Effects of deepwater trawling on the macro-invertebrate assemblages of seamounts on the Chatham Rise, New Zealand. Deep Sea Res. I 56, 1540-1554. doi: 10.1016/j.dsr.2009.04.015
CrossRef Full Text | Google Scholar
Clark, M., Tracey, D., Anderson, O., and Parker, S. (2014). Pilot Ecologische Risicobeoordeling voor Beschermde Koralen. Wellington: National Institute of Water and Atmospheric Research, 32.
Google Scholar
Clark, M. R., Bowden, D. A., Rowden, A. A., and Stewart, R. (2019). Little evidence of benthic community resilience to bottom trawling on seamounts after 15 years. Front. Mar. Sci. 6:63. doi: 10.3389/fmars.2019.00063
CrossRef Full Text | Google Scholar
Coleman, D. C. (1991). Koolstofisotoop Technieken. Cambridge, MA: Academic Press.
Google Scholar
Consalvey, M., MacKay, K., and Tracey, D. (2006). Information review for protected deep-sea coral species in the New Zealand region. NIWA Client Report. WLG2006-85. 60. Auckland: NIWA.
Google Scholar
Druffel, E. R. M., Griffin, S., Witter, A., Nelson, E., Southon, J., Kashgarian, M., et al. (1995). Gerardia: borstelkegels van de diepzee? Geochim. Cosmochim. Acta 59, 5031-5036. doi: 10.1016/0016-7037(95)00373-8
CrossRef Full Text | Google Scholar
Druffel, E. R. M., King, L. L., Belastock, R. A., and Buesseller, K. O. (1990). Groeisnelheid van een diepzeekoraal aan de hand van 210Pb en andere isotopen. Geochim. Cosmochim. Acta 54, 1493-1500.
Google Scholar
Fallon, S. J., Fifield, L. K., and Chappell, J. M. (2010). Het volgende hoofdstuk in radiokoolstofdatering aan de Australian National University: statusrapport over het eentraps AMS. Nuclear Instr. Methods Phys. Res. Sec. B 268, 898-901. doi: 10.1016/j.nimb.2009.10.059
CrossRef Full Text | Google Scholar
Freeman, D., and Cryer, M. (2019). “Current management measures and threats,” in The State of Knowledge of Deep-Sea Corals in the New Zealand Region, Chap. 9, eds D. M. Tracey and F. Hjorvarsdottir (Auckland: NIWA Science and Technology Series).
Google Scholar
Gammon, M. J., Tracey, D. M., Marriott, P. M., Cummings, V. J., and Davy, S. K. (2018). De fysiologische respons van het diepzeekoraal Solenosmilia variabilis op oceaanverzuring. PEERJ 6:e5236. doi: 10.7717/peerj.5236
PubMed Abstract | CrossRef Full Text | Google Scholar
Griffin, S., and Druffel, E. M. (1989). Sources of carbon to deep-sea corals. Radiocarbon 31, 533-543. doi: 10.1017/s0033822200012121
CrossRef Full Text | Google Scholar
Hitt, N. T., Sinclair, D. J., Fallon, S. J., Neil, H. L., Tracey, D. M., Komugabe-Dixson, A., et al. (2020). Groei en levensduur van Nieuw-Zeelandse zwarte koralen. Deep Sea Res. I 2020:103298. doi: 10.1016/j.dsr.2020.103298
CrossRef Full Text | Google Scholar
Komugabe, A. F., Fallon, S. J., Thresher, R. E., and Eggins, S. M. (2014). Modern Tasman Sea surface reservoir ages from deep-sea black corals. Deep-Sea Res. 99, 207-212. doi: 10.1016/j.dsr2.2013.05.024
CrossRef Full Text | Google Scholar
Komugabe-Dixson, A. F., Fallon, S. J., Eggins, S. M., and Thresher, R. E. (2016). Radiokoolstof bewijs voor mid-late Holocene veranderingen in zuidwestelijke Stille Oceaan circulatie. Paleoceanography 31, 971-985. doi: 10.1002/2016PA002929
CrossRef Full Text | Google Scholar
Love, M. S., Yoklavich, M. M., Black, B. A., and Andrews, A. H. (2007). Age of black coral (Antipathes dendrochristos) colonies, with notes on associated invertebrate species. Bull. Mar. Sci. 80, 391-400.
Google Scholar
Matsumoto, K. (2007). Op radiokoolstof gebaseerde circulatieleeftijd van de wereldoceanen. J. Geophys. Res. 112:C09004. doi: 10.1029/2007JC004095
CrossRef Full Text | Google Scholar
Mortensen, P. B., and Buhl-Mortensen, L. (2005). Morphology and growth of the deep-water gorgonians Primnoa resedaeformis and Paragorgia arborea. Mar. Biol. 147, 775-788. doi: 10.1007/s00227-005-1604-y
CrossRef Full Text | Google Scholar
Noé, S. U., Lembke-Jene, L., and Chr Dullo, W. (2008). Varying growth rates in bamboo corals: sclerochronology and radiocarbon dating of a mid-Holocene deep-water gorgonian skeleton (Keratosis sp.: Octocorallia) van Chatham Rise (Nieuw-Zeeland). Facies 54, 151-166. doi: 10.1007/s10347-007-0129-x
CrossRef Full Text | Google Scholar
Pinkerton, M. H. (2016). Ocean Colour Satellite Observations of Phytoplankton in the New Zealand EEZ, 1997-2016. Ministerie voor Milieu. Wellington: NIWA.
Google Scholar
Prouty, N. G., Fisher, C. R., Demopoulous, A. W. J., and Druffel, E. R. M. (2016). Growth rates and ages of dee-sea corals impacted by the Deepwater Horizon oil spill. Deep-Sea Res. II 129, 196-212. doi: 10.1016/j.dsr2.2014.10.021
CrossRef Full Text | Google Scholar
Prouty, N. G., Roark, E. B., Buster, N. A., and Ross, S. W. (2011). Growth rate and age distribution of deep-sea black corals in the Gulf of Mexico. Mar. Ecol. Prog. Ser. 423, 101-115. doi: 10.3354/meps08953
CrossRef Full Text | Google Scholar
Reimer, P. J., Bard, E., Bayliss, A., Beck, J. W., Blackwell, P. G., Bronk Ramsey, C., et al. (2013). IntCal13 en Marine13 radiokoolstof leeftijdkalibratiecurves 0-50.000 jaar cal BP. Radiocarbon 55, 1869-1887. doi: 10.2458/azu_js_rc.55.16947
PubMed Abstract | CrossRef Full Text | Google Scholar
Reimer, P. J., and Reimer, R. W. (2001). A marine reservoir correction database and on-line interface. Radiocarbon 43, 461-463. doi: 10.1017/s0033822200038339
CrossRef Full Text | Google Scholar
Roark, E. B., Guilderson, T. P., Dunbar, R. B., Fallon, S. J., and Mucclarone, D. A. (2009). Extreme levensduur bij proteïnerijke diepzeekoralen. Proc. Natl. Acad. Sci. U.S.A. 106, 5204-5208. doi: 10.1073/pnas.0810875106
PubMed Abstract | CrossRef Full Text | Google Scholar
Roark, E.B., Guilderson, T.P., Dunbar, R.B., and Ingram, B.L. (2006). Op radiokoolstof gebaseerde leeftijden en groeisnelheden: Hawaiian deepsea corals. Mar. Ecol. Progr. Ser. 327, 1-37.
Google Scholar
Sánchez, J., and Rowden, A. (2006). “Octocoral diversity on New Zealand Seamounts,” in Proceedings of the Presented at the 10th International Coral Reef Symposium, Okinawa.
Google Scholar
Sherwood, O.A., and Edinger, E.N. (2009). Ages and growth rates of some deep-sea gorgonian and antipatharian corals of Newfoundland and Labrador. Can. J. Fish. Aquatic Sci. 66, 142-152. doi: 10.1139/f08-195
CrossRef Full Text | Google Scholar
Sherwood, O. A., Scott, D. B., Risk, M. J., and Guilderson, T. P. (2005). Radiocarbon evidence for annual growth rings in the deep-sea octocoral Primnoa resedaeformis. Mar. Ecol. Prog. Ser. 301, 129-134. doi: 10.3354/meps301129
CrossRef Full Text | Google Scholar
Sikes, E., Burgess, S., Grandpre, R., and Guilderson, T. (2008). Assessing modern deep-water ages in the New Zealand region using deep-water corals. Deep Sea Res. I 55, 38-49. doi: 10.1016/j.dsr.2007.10.004
CrossRef Full Text | Google Scholar
Stuiver, M., and Polach, H. A. (1977). Discussie rapportage van 14C gegevens. Radiocarbon 19, 355-363. doi: 10.1017/S0033822200003672
CrossRef Full Text | Google Scholar
Thresher, R., Rintoul, S. R., Koslow, J. A., Weidman, C., Adkins, J., and Proctor, C. (2004). Oceanic evidence of climate change in southern Australia over the last three centuries. Geophys. Res. Lett. 31:l07212.
Google Scholar
Thresher, R. E., MacRae, C. M., Wilson, N. C., and Gurney, R. (2007). Environmental effects on the skelet composition of deep-water gorgonians (Keratoisis spp.; Isididae). Bull. Mar. Sci. 8, 409-422.
Google Scholar
Tracey, D., Bostock, H., and Shaffer, M. (2018). Ageing Methods for Protected Deep-Sea Corals: A Review and Recommendation for an Ageing Study. DOC Contract 4527 GMC – Leeftijd en Groei van koraal (POP2017-07). NIWA Client Report WN2018035. Auckland: NIWA, 40.
Google Scholar
Tracey, D., Neil, H., Gordon, D., and O’Shea, S. (2003). Chronicles of the deep: ageing deep-sea corals in New Zealand waters. NIWA Water Atmosph. 11, 22-24.
Google Scholar
Tracey, D. M., and Hjorvarsdottir, F. (eds) (2019). “The state of knowledge of deep-sea corals in the new zealand region,” in NIWA Science and Technology Series Number 84, (Auckland: NIWA),140.
Google Scholar
Tracey, D. M., Neil, H., Marriott, P., Andrews, A. H., Cailliet, G. M., and Sanchez, J. A. (2007). Age and growth of two genera of deep-sea bamboo corals (family Isididae) in New Zealand waters. Bull. Mar. Sci. 81, 393-408.
Google Scholar
Williams, A., Schlacher, T. A., Rowden, A. A., Althaus, F., Clark, M. R., Bowden, D. A., et al. (2010). Seamount megabenthic assemblages niet te herstellen van trawlvisserij impacts. Mar. Ecol. 31, 183-199. doi: 10.1111/j.1439-0485.2010.00385.x
CrossRef Full Text | Google Scholar
Williams, B., and Grottoli, A. G. (2010). Stabiele stikstof en koolstof isotopen (δ15N en δ13C) variabiliteit in ondiepe tropische Pacific zacht koraal en zwart koraal taxa en implicaties voor paleoceanografische reconstructies. Geochim. Cosmochim. Acta 74, 5280-5288. doi: 10.1016/j.gca.2010.06.026
CrossRef Full Text | Google Scholar
Williams, B., Risk, M. J., Ross, S. W., and Sulak, K. J. (2007). Stabiele isotopengegevens van diep-water antipathariërs: 400-jaar gegevens van de zuidoostkust van de Verenigde Staten van Amerika. Bull. Mar. Sci. 81, 437-447.
Google Scholar
Leave a Reply