Frontiers in Marine Science
はじめに
深海珊瑚は非常に多様な海洋生物のグループであり、ニュージーランド地域は深海珊瑚のホットスポットである(Sánchez and Rowden, 2006; Cairns et al, 2009). いくつかの研究では、南太平洋地域における深海の大型無脊椎動物群(サンゴを含む)に対する底引き網漁の影響を調査している(Althaus他、2009;ClarkとRowden、2009;Clark他、2010、2019;Williams他、2010;Anderson他、2014)。 これらは、底引き網漁が、成長が遅く壊れやすい深海のサンゴのコロニーや群集に大きな影響を与え、回復する能力がほとんどないことを実証しています。 気候変動から予測される海洋温暖化や海洋酸性化などの環境影響も、Anthony and Marshall(2009)において深海サンゴが繁栄するには暖かくなりすぎる生息地や、pHや炭酸イオンが減少し、炭酸ベースの骨格を持つ深海サンゴに影響を与えると考えられる(Gammon et al, 2018)ため、深海の健全性が脅かされることが予想されている。 将来のリスク評価や海洋資源管理により良い情報を提供するために、深海サンゴの年齢と成長を理解することは、人為的な妨害やその他の自然被害後のサンゴ再生時期や回復可能性を決定する鍵となる。
ニュージーランド地域では、深海サンゴの分類、異なる種の分布とその環境選好、底生生態系に対する深海サンゴの重要性に関するかなりの研究が行われてきた。 最近、ニュージーランド地域の深海サンゴに関する理解と知識のレビューが発表された(Tracey and Hjorvarsdottir, 2019)。 このレビューでは、多くの属や種について年齢や成長のデータが限られているなど、深海珊瑚の理解における大きなギャップについても概説されている。 本研究では、保護されている深海の黒サンゴAntipatharian Bathypathes patulaの年齢と成長速度を評価する。 このサンゴの年齢と成長速度は、この地域ではこれまで研究されていない。 この種の選択は、リスクアセスメントの優先順位リスト(Clark et al., 2014)と文献レビュー(Tracey et al., 2018)に基づき、サンプル(場所と総数)の入手可能性と、ニュージーランドで行われている他の補完的な黒サンゴ研究(Hitt et al., 2020)と相まって行われた。
Tracey et al. (2018)は深海サンゴ種の老化に利用されている現在の方法について概要を示した。 各手法の利点と欠点が議論された。 深海サンゴの年齢と成長の測定に成功した主な方法は、(1)線状成長または表面拡張のin situ測定または水槽内実験などの直接観察、ポリプ付加率、石灰化率の推定(例, using the buoyant weight technique)、(2)骨格成長帯やバンドの列挙(sclerochronology)、(3)放射性物質分析
深海サンゴは、成長に時間がかかるため、生きた生検体の入手や飼育が難しく、直接観察およびin situや水槽内での測定は非常に難しい(Gammon et al.2018; Tracey et al.2018). 成長帯の列挙、スクレロクロノロジーは、骨格断面で観察される帯状の成長パターンのコントラストが高い深海性サンゴ種に最適である。 しかし、観察された成長帯のカウントの年代的妥当性を検証するために、独立した検証方法を用いることが不可欠である (Andrews et al., 2002)。 この方法は、400 年前のタケノコサンゴ Keratoisis sp.を含むゴルゴニアン八重サンゴの年齢と成長率データを決定するのに成功した。 100 歳のウミウシ Primnoa spp.、60 歳のタケノコサンゴ Lepidisis sp. (Thresher et al., 2004, 2007; Mortensen and Buhl-Mortensen, 2005; Sherwood et al., 2005; Tracey et al., 2007; Sherwood and Edinger, 2009)。 黒サンゴは、成長域や個体数を用いた年代測定にも成功しており、Antipathes dendrochristos と Leiopathes glaberrima では、それぞれ 150 歳から 480 歳の年代を記録した(Love et al.2007; Williams et al.2007)。 黒サンゴの Stauropathes arctica の研究では、ゾーンカウントが 55-58 となり(Sherwood and Edinger, 2009)、著者らは放射性炭素年代測定により、これらの年齢が 55 年と 82 年に制約されると述べている。 成長帯の列挙による黒サンゴの成長率は、8〜140μm/yrの低い放射状成長を明らかにした(Love et al., 2007; Prouty et al., 2011)
成長帯の列挙法には、潜在的限界がある(Tracey et al., 2018の要約を参照)。 多くのサンゴは、骨格全体に明確な規則的なゾーン構造を示さないか、明確なゾーン構造は骨格の一部にしか現れないことがある。 存在する場合でも、ゾーン構造は可変であるか、異なる解釈の可能性を示すことがある。 例えば、タケノコガイ科のKeratoisis sp.とLepidisis sp.の基部と先端部のカウントは、鉛210(210Pb)放射計法で推定した年代よりも高かった(Tracey et al.、2007年)。 光学顕微鏡で観察されたゾーンは2年ごとの周期性を持っており、SEMで観察された結節点のゾーンは月の周期性などの環境イベントを表している可能性があると仮定された。 月の周期は強い潮汐と関連している可能性があり、その結果、無脊椎濾過摂食生物にとって餌の利用可能性が高まり、体細胞成長のパルスが発生する可能性がある。 ゴルゴニアン バブルガムコーラル(Paragorgia spp.など)やレッドプレシャスコーラル(Corallium sp.など)は、帯状パターンが複雑で定義しにくいため、ゾーンカウントではうまく熟成できない(Griffin and Druffel, 1989; Andrews et al.)。 2005)。
深海サンゴに用いられる最も一般的な放射性測定法は、放射性炭素(14C)同位体を用いた年代測定である(Druffelら、1995; Sherwoodら、2005; Traceyら、2018)。 この方法は、大気中のごくわずかな自然炭素が14Cの形で放射性であり、サンゴの骨格が炭酸カルシウムやタンパク質、キチンを形成する際に取り込まれることを利用している(Adkins et al., 2002; Tracey et al., 2003; Consalvey et al., 2006)。 放射性炭素の半減期は5,730年と知られているので、この方法は高価ではあるが、5万年前以下の標本の年代を確実に測定することができる。それ以上では、14Cの活性が低くなりすぎて検出できない(Coleman, 1991)。 1950年代から60年代にかけて、核実験によって大気中の14Cが急増した。この「爆弾炭素」も年代を校正する道具として使用でき、より新しい試料の基準点となる(Coleman, 1991; Druffel et al, 1995; Tracey et al, 2003; Roark et al.,)。 2009; Sherwood and Edinger, 2009)。
放射性炭素年代測定は、石サンゴ、黒サンゴ、ゴルゴニアン八重サンゴなど、いくつかのサンゴ種でうまく使用されている(Druffelら, 1990; Roarkら, 2006; Carreiro-Silva et al, 2013; Proutyら, 2016; Hittら, 2020)。 しかし、この方法には限界もある。 海洋循環の結果、海洋の水の放射性炭素年代は大気よりかなり古く、いわゆる「海洋リザーバー年代」と呼ばれています(Tracey et al., 2003; Consalvey et al., 2006; Roark et al., 2006; Noé et al., 2008)。 リザーバー年齢の誤差は、サンゴの年齢推定に大きな影響を与える可能性があります。 リザーバー年代の補正には、周辺海域の知識が必要であり、水中の放射性炭素の比率が一定であることを仮定しているが、これらのレベルは、海洋内の混合、局所的な湧昇現象、生産性ブルームによって変化することがあり、これらはすべて、炭素循環変換と14Cのサンゴ骨格組成を変化させることができる(Tracey et al, 2018)。
Traceyら(2018)のレビューに基づいて、我々はB. patulaの2つの年代測定方法を使用することにしました; 骨格成長バンドのカウントと放射性炭素年代測定法(14C)です。 これらの方法を適用することで、ニュージーランドと世界の他の過去のサンゴの年代測定研究と比較することができます。 サンプルの選択、材料のマイクロミル加工、切片作成方法、ゾーンカウントと放射性炭素年代測定の解釈について紹介する。
材料と方法
試料の選択
本研究で用いた試料は、ニュージーランド国立水・大気研究所(NIWA)の国立無脊椎動物コレクション(NIC)に保存されている試料から得られたものであった。
サンプルは、ベイ・オブ・プレンティ(ニュージーランド北島北東部)とチャタムライズ(ニュージーランド東部)の2地域から選び、異なる地域間の成長速度の結果比較と評価、また同じ地域の他の深海黒サンゴ属、特にAntipathella、Leiopathes、Antipathesとも比較した(Hitt et al., 2020)。
海流は、餌の安定した流れや幼生の分散をもたらし、深海珊瑚に影響を与える。 その結果、これらの海流は、餌の入手可能性や深海珊瑚の分布にも影響を与える。 深海珊瑚は、その上を流れる底層水流の影響を最も受け、堆積物の分布に影響を与えますが、表層水流の影響も受け、基礎生産力に影響を与え、海底の深海珊瑚に降り注ぐ餌の供給量に影響を与えるのです。 ニュージーランドは、北から流れ込む暖かく栄養分の少ない亜熱帯表層水と、南から流れ込む冷たく栄養分の多い亜南極表層水の交差点に位置している。 この地域の表層、サーモクライン、中間水域、深層水域の水塊と海流に関する現在の理解は、最近Chiswellら(2015)がレビューしている。 本研究によるサンゴの海洋学的設定は以下の通りである
ベイ・オブ・プレンティは、東オーストラリア海流から供給され、西から流入して東オークランド海流を形成する、温暖で低栄養の亜熱帯表流水で覆われている。 チャタム海峡は、暖かく低栄養の亜熱帯海水(STW)と南氷洋からの冷たく高栄養の亜寒帯海水(SAW)が出会う地点で、亜熱帯前線(STF)である。 STF での STW と SAW の水塊の混合により、チャタム海峡は非常に高い基礎生産性の地帯となり、これらのサンゴに重要な食料源とそれゆえの炭素供給を提供している。クロロフィル a 濃度は、中央太平洋の 0.03-0.3 mg/m3 に対して、チャタム海峡とプレンティ湾では 0.3-0.48 mg/m3 に及ぶ (Pinkerton, 2016)。 水深500~1500mでは、冷温(3.5~10℃)、低塩分(34.4~34.6)、高酸素(200~250μmol/kg)、高栄養(硝酸15~36μmol/kg)の南極中間水(AAIW)が北と南からこの地域に流れ込む(Chiswell et al, 2015)。
チャタムライズからのサンゴは、この領域からの現代の14Cリザーバー年齢のデータ(Sikesら、2008)があることから選ばれた。
サンプルはニュージーランド漁業省(FNZ)、第一次産業省(MPI)オブザーバー、NIWA研究者によって収集された既存の標本から選択し、(NIC)に保管されている。 B. patulaの分布データをプロットすることで、コロニーサイトの選定と標本数の決定に役立てた(図1)。 ベイ・オブ・プレンティから2つの標本(NIWA24190とNIWA85940)が放射性炭素年代測定とゾーンカウントのための薄切片として選ばれた。 残りの標本はチャタムライズから選んだ。 ニュージーランド地域でBathypathes patulaが標本化された場所。 赤丸はニュージーランド国立無脊椎動物コレクション(NIC)に保管されているBathypathes patulaのサンゴ標本の個々の採取地。 灰色の線は水深 200m の等高線。 黒線はニュージーランドの排他的経済水域(EEZ)の範囲を示す。
2018年末、黒サンゴ分類学の客員専門家(ジェレミー・ホロウィッツ、ジェームズクック大学)がNICのいくつかのサンプルの同定を見直した。 この過程で、今回の年齢推定調査のために選定され、Bathypathes patulaと想定された黒サンゴのコロニーの1つが、同定を見直された。 NIWA24190は、Bathypathes cf. confertaと同定されました。 この論文では、このサンプルを Bathypathes cf. conferta と表記しています(図2)。 この標本の分類学的再検討は現在進行中である(Opresko pers comm)。 分布図(図1)には、B. cf. confertaの他の例も含まれているかもしれない。 Bathypathes cf. conferta. NIWA24190。 解析のために試料を採取した場所を示す標本。 (A)は成長した先端部、(B)は基底部。 ニュージーランド、ベイ・オブ・プレンティ産の標本。 スケールバーは35cm定規。
材料の準備
適切なサンプルを特定したら、サンゴの主茎の成長先端と基部から小さな(10mm以下の)切片を切り出した。 また、サンプルの多くは採取時に破損していたため、線状成長率を確実に推定するために、主茎に沿ってさらに切片を採取しました(図2-5)。 一つは放射性炭素年代測定用、もう一つはゾーンカウントを得るための薄切片作成用です。

Figure 3. Bathypathes patula NIWA85940。 解析のために試料を採取した場所を示す標本。 (A)は成長した先端部、(C)は基部、(B)は中間部である。 ニュージーランド、ベイ・オブ・プレンティ産の標本。 スケールバーは35cm定規。

Figure 4. Bathypathes patula NIWA49468。 解析のために試料を採取した場所を示す標本。 (A)は成長先端部、(D)は基底部、(B,C)は中間部であり、採集時に標本が破損し、この2箇所の間で主枝幹の長さが不明になっている可能性がある。 ニュージーランド、チャタムライズからの標本。 スケールバーは35cm定規。

Figure 5. Bathypathes patula NIWA47911. 分析のためにサンプルを採取した場所を示す標本。 (A)は成長した先端部、(F)は基底部、(B-E)は放射性炭素爆弾のシグナルを探すために採取した中間試料部位です。 ニュージーランド、チャタムライズからの標本。
Micro-Milling of Material
これまでの研究から、黒サンゴの切片は毛細管現象によって多孔質の母体から樹脂を吸い上げやすいことが指摘されており、この研究の切片は樹脂でベースプレートに接着して切削することができなかった。
NIWAのNew WaveTMマイクロミルに0.5mmのBrassler H2.11.006ミリングバリを取り付け、基部、中部、先端部の端と芯から粉末の放射性炭素試料を採取しました。 エッジサンプルは切片の外側250ミクロンの材料から、コアサンプルは原基の半径750ミクロン以内の材料から得られたものである。 1試料あたり少なくとも1.5mgの試料を抽出するよう努めた。
Thin Sectioning Method and Zone Counting Protocols
Coral stem sections were embedded in clear two-part epoxy resin and sectioning with a diamond-wafering saw at NIWA. 切片は片面を研磨し、研磨面を下にしてガラス製の顕微鏡用スライドに取り付け、透過光で成長帯構造を見るのに最適の厚さになるまで研磨した。 この種の切片に最適な厚さは約250-300μmで、薄切片作成の標準的な手順に従った (Andrews et al., 2002; Mortensen and Buhl-Mortensen, 2005; Tracey et al., 2007)。 毎年形成されると想定される周期的な成長バンドまたは増分を数えることは、NIWAでの薄切片作成後に行われた。
ゾーンカウントを行うために、主枝セクションの薄切片作成を透過光のある複合顕微鏡で観察した。 また、Sherwood and Edinger (2009)の方法に従って、ゾーン解釈の補助として、紫外線を使用して切片を観察した。
Radiocarbon Dating Method
分析に先立ち、試料は黒皮と内石を除去し、結果を変える可能性のある若い汚染物質14Cを除去した (Adkins et al., 2002)。 粉末サンゴ試料を<1 mlの0.1 M HCLで処理した。 その後、酸をピペッティングで除去し、試料をmilliQ水で3回洗浄した。 その後、試料をドライアイスで凍結し、凍結乾燥した。 洗浄したサンゴ試料約1mgを外径6mmの石英管に秤量し、約60mgのCuOと銀杯を加えた。 石英管は<3e-3 torr (<0.4 Pa)まで排気し、トーチで密封した。 その後、チューブを900℃で6時間焼成し、CO2を発生させた。 得られたCO2を精製し、水素の存在下、粉末状のFeを触媒としてグラファイトに変換した。 その後、グラファイトはANUのシングルステージ加速器質量分析計で測定された(Fallon et al., 2010)。 すべてのデータはオンライン AMS δ13C で補正し、シュウ酸 I で規格化し、サンゴ試料と同じ方法で処理した 14C フリー石炭を用いてバックグラウンド減算を行った。 データはStuiver and Polach (1977)の勧告に従って表示した。
結果
ゾーンカウントのための薄切片
これらの種の骨格標本で観察されたゾーン構造パターンは非常に複雑で、多くの方法で解釈することが可能であった。 径方向成長数については、準備した切片を注意深く観察し、原基から外縁までの明確なゾーン構造を最もよく示す軸を使用してゾーンカウントを求めた。 B. patulaの枝の半径方向の成長は適度に規則的で、他の黒サンゴ種でこれまで観察されてきた、枝の特定の側に厚いゾーンを形成する顕著な偏りはなかった。 また、成長率の計算に用いた半径は、準備した切片で観察された最小半径と最大半径の平均であり、報告された半径成長率はそのサンゴの平均半径成長率である。
サンゴ骨格には、低倍率(10-20倍)で観察すると、暗色のゾーンと明るいゾーンが交互に並ぶ粗いバンド模様(図6)が大きく観察される。 また、高倍率(100〜200倍)で観察すると、適度に規則的な細かい帯状模様も確認できる(図7)。 ここでは、粗く形成された広いゾーンと、細く形成された微細なゾーンの両方を数えるという、2つの読み取りプロトコルを適用した(表1)。 ここで紹介するゾーンカウントは、すべて1人のリーダー(Marriott)によって行われたものである。 ゾーン解釈プロトコルを開発する過程で、開発中のプロトコルの様々な反復を適用して多くの読み取りが行われた。 ここに示した2つのプロトコルのゾーンカウントは、それぞれセクション全体にわたる複数のカウントから得られたものである。 サンゴNIWA47911の底部断面。観察されたゾーン構造の粗い色帯の解釈を示す。 標本には赤い点で示した54のゾーンがある。 挿入した破線の長方形は図7で撮影した基底部の中央部を示す。 観察されたゾーン構造のファインスケール解釈プロトコルを示すサンゴNIWA47911の底部中央部分。 この標本では、この断面内側の領域に黒い点で示された42のゾーンがあり、断面全体では211のゾーンがあった。

Table 1.
試験片の最も若い部分である先端または先端付近の領域では、4〜7(粗いゾーン数)、10〜19(細かいゾーン数)であった。 主枝の中間領域では、カウントは55から60(粗帯カウント)、370から374(細帯カウント)であった。 コロニー底部では、37から193(粗大ゾーン数)、78から481(細粒ゾーン数)であった。
Radiocarbon Analysis
表2に示した粉砕試料をANUに送り、放射性炭素年代測定を実施しました。 17個のサンゴ試料の結果は放射性炭素測定結果(Table 3)に示されています。 3つの試料(Sample 47911-F2; 24190-B1; 85940-B1)は実験室で失敗し、複製を試みるには十分な材料が残っていませんでした。 残りのサンゴの放射性炭素の結果は、OxCal4.3 (Bronk Ramsey, 2001)、Marine13曲線、ニュージーランドローカルのdeltaR (radiocarbon reservoir offset) -18 ± 36 (Reimer and Reimer, 2001) を用いて暦年代に較正されました。 原爆投下前のサンゴ試料の年齢範囲分布を表 3 に示す。 なお、本研究で使用したサンゴ標本はすべて生きたまま捕獲されたものであり、推定年齢は標本の寿命ではなく、捕獲時の年齢である。 この種の寿命は、ここで示した推定値よりも長くなることが予想されます。 サンゴ 85940 は約 380±50 歳、サンゴ 49468 は約 310±70 歳、サンゴ 47911 は約 120±50 歳でした(表 3)。 また、サンゴの外縁部と先端部には、85940-B2と85940-C2を除き、放射性炭素値F14C >1が含まれており(表3)、これらのサンゴは採取時に生きていて表層水の炭素を取り込んでいたことが分かります。 85940-B2と85940-C2は外縁部の放射性炭素値がF14C <1 (Table 3) であり、骨格の一部が活発に物質を取り込んでいなかったことがわかります。 これは、サンゴコロニーの基部は成長を止めるが、上部は成長を続けるという現象が以前から観察されている(Komugabe-Dixson et al.、2016)
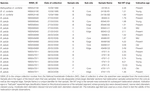
Table 2. 放射性炭素年代測定のために抽出されたサンプル.

Table 3.
Growth Rate Estimates
成長率推定は放射性炭素年代測定結果に基づいて行われました。 成長率はサンプリングポイント間の物理的距離と、関連するサンプルサイトでの放射性炭素年代をもとに算出されました。 放射性炭素分析中にいくつかの試料が不合格となったため、限られた成長率の推定値しか作成できませんでした。 径方向成長率(コロニーの枝が時間とともに径方向に太くなる率)は、コアからサンプルサイトの端までの成長の暦年数を枝セグメントの平均半径で割って推定した。 半径方向の成長速度は11.1〜35.7μm/yrであった。 線形成長率(コロニーの枝が時間とともに縦方向に伸びる率)は、コアサンプルの較正暦年代の差を2つのサンプルサイト間の直線距離で割って推定した。 その結果、5.2mm/yrと9.6mm/yrの2つの直線成長率が得られた(表4)。

Table 4. Bathypathes patula growth rate estimates.
Discussion
Bathypathes patula Age and Growth Estimates
今回の放射性炭素の結果からB. patulaは長寿種であり、385年を超える年代があることがわかりました。 また、放射性炭素年代測定結果より、ゆっくりとした線形成長率(5.2-9.6mm/yr)と放射状成長率(11.1-35.7μm/yr)が得られた。
ゾーンカウントプロトコルはいずれも放射性炭素年代推定値と相関が認められなかった。 微細なゾーンカウントは14C年代と最も近い相関を示し、ゾーンカウントと14C年代を比較した場合、長寿であることを全体的に支持する結果となった。 放射性炭素年代データとファインスケールゾーンカウンティングデータの間の年代推定値は、112年から385年の放射性炭素年代を持つ枝セクションにおいて、日常的に100年以上の相違が見られた(表1、3)。 2つの方法による比較可能な試料からの推定年代の差は負と正であり、ゾーンカウントを用いた場合、それに比例して年代が過小になったり過大になったりすることはなかった(表1)。
標本NIWA-85940の端部放射性炭素の結果は、主茎の基底部(C2)が収穫前293年に、中間部(B2)が収穫前88年に骨格物質の堆積を停止したことを示唆した。 このことは、これらの枝幹の部分は、生きたポリプを持つ側線小節に隣接していることが観察されたにもかかわらず、である。 これらのセクションの光学的外観は、その外側の縁でゾーン堆積が停止したことを示すものではありませんでした。 また、サンゴの枝の表面は比較的きれいで、新しい骨格が付着していないサンゴの枝に見られるような、表面的な浸食や変色は見られず、サンゴの枝の表面はきれいなままでした。 このことは、もし枝の先端が成長し続ける一方で、枝の茎が骨格の蓄積を止めることができるならば、この種のサンゴの基底部切片からの年齢推定にゾーンカウント法のプロトコルが信頼できる形で利用されないことを示唆するものである。 これらのサンプルは25mm間隔で配置された。 これは、サンゴの骨格に爆弾放射性炭素が出現する位置を特定するための試みです。 これは、サンゴの骨格に爆弾放射性炭素が出現した場所を特定する試みで、これにより、コロニーの枝の先端がより成長し、線形成長速度を正確に評価することができます。 また、このコロニーと同じ水塊から採取された他のコロニーに適用する14C補正を設定するための正確な基準点を得ることができます。 残念ながら、これらのサンプルはすべて原爆投下後であることが判明し、この作業は完了しませんでした。 今後、この枝幹をさらにさかのぼって試料を採取することで、このタイポイントや現代の線形成長率を確定することができると思われます。 私たちの研究では、5.2-9.6mm/yrの線形成長率を測定しました。最も速い成長率を仮定すると、骨格中の爆弾放射性炭素の上昇を確実に観察するためには、成長先端から主枝幹の700mm後方までサンプルを採取する必要があると考えられます。
今回の放射性炭素測定では、成長枝の先端と主枝表面の試料でF14C >1を示し、これらの試料が現代の原爆後レベルを示していることが示された。 海水混合層より下に原爆後の炭素が存在することは、B. patula黒サンゴが最近輸出された表層水のPOCを使用していることを示す。 この深度では、POCは爆弾14Cでタグ付けされた唯一の炭素源です。 サンゴが生息している環境水の溶存無機炭素(DIC)は、14C値がはるかに低い(Matsumoto, 2007; Alves et al.、2018)。 爆弾14Cを経験していない海域で生育するサンゴから爆弾14Cを検出することは、黒サンゴが表面由来のPOCから炭素を得ていることを補強する(Roark et al., 2009; Sherwood and Edinger, 2009; Williams and Grottoli, 2010; Prouty et al.、2011)。 このことは、石サンゴのように深海のリザーバー効果をモデル化する必要がないことを意味する。石サンゴは、骨格の炭素を中間水域のDICから得て、それを浴びることができる。
最近のニュージーランドおよび世界の黒サンゴ年齢データとの比較
本プロジェクトによる放射性炭素由来の年齢推定値と成長率は、ニュージーランド地域の主要な深海黒サンゴ(Antipatharia)について最近行われた他の多くの年齢および成長研究の結果と同様である(Table 5)。 これらの推定年齢と成長率は、B. patulaは、Leiopathes、Antipathes、Antipathellaのコロニーなど、他のサンゴ属と同様、2600年まで生き、1-143μm/yrのオーダーで成長する(Hittら、2020年)。 今回の結果は、ニュージーランドの黒サンゴがこの地域で最も成長の遅い海洋動物であり、一部の種は極端な長寿を示すことをさらに裏付けるものとなった。

Table 5. ニュージーランド産黒サンゴ(Antipatharia)の放射性炭素年代推定研究
ニュージーランドで年代測定されたB. patulaや他の黒サンゴの結果は、これらのサンゴが生息する水塊と深さに関して、成長率や寿命の明確な傾向を示していない。 Chatham Rise と Bay of Plenty のサンゴは、750~1269 m のほぼ同じ水深で採取されたものが多く、Chatham Rise の B. patula は 1120~1269 m の深い場所で採取されたものでした(表 1、表 5)。 Chatham Riseの標本(NIWA-49468とNIWA47911)の半径方向の成長速度は20-36μm/yrで、Bay of Plentyの標本(NIWA-85940)の11μm/yrより高い。 この2つの地域のB. patula標本では3つの成長率しか得ることができなかったので、成長率の地域差について仮定することはできない。 Chatham Rise の黒サンゴの成長速度データを見ると、Antipathes sp.の標本が 91-143 μm/yrと最も速く、Leiopathes secunda の標本が 5-15 μm/yrと最も遅い。 ベイ・オブ・プレンティ産黒サンゴのBathypathes patula、Leiopathes secunda、Leiopathes sp.は5-40μm/yrと中位から低い径方向成長速度であった。 一方、フィヨルドの浅海域の黒サンゴAntipathella fiordensisは比較的高い半径方向成長率(50-57μm/yr)を示すが、これが種によるものかフィヨルドの環境条件によるものか、あるいは両者の混合なのかは不明である
この研究はまた、黒サンゴが最も成長の遅い深海サンゴであるとした世界の研究とも一致している。 Sherwood and Edinger (2009) は、黒サンゴ Stauropathes arctica の年齢を最大82暦年まで推定しています。 Prouty et al. (2011)は、捕獲した Leiopathes sp.のコロニーについて、放射性炭素を用いて 2100 年まで の生年月日を推定している。 Carreiro-Silvaら(2013)は、アゾレス諸島のLeiopathes sp.のコロニーを2320歳と推定している。 ハワイのLeiopathes annosahasの標本は、4000年を超える寿命に達する可能性を示した(Roark et al.、2009)。
この新しい年齢と成長率は、深海ブラックサンゴB. patulaの繊細な年齢を強調し、本研究では385年を超える年齢に到達し、遅い線形および放射状の成長速度を示す長寿種であることが示された。 この結果は、攪乱・除去後の回復力が低く、回復に時間がかかる可能性があることを示しています。 この研究は、ニュージーランドのサンゴ管理の枠組み(Freeman and Cryer, 2019)における今後のリスク評価と管理戦略に活用される。 成長が遅く、極めて長寿であることから、これらのサンゴは、漁業や採掘などの海底攪乱に対して非常に脆弱であり、人為的な影響から回復するのに数世紀かかる可能性があることがさらに浮き彫りになった(Hitt et al, 2020)。
結論
ニュージーランドの2つの地域、ベイ・オブ・プレンティとチャタムライズから採取した深海クロサンゴ B. patula の年齢と成長速度を、成長環計数と放射性炭素年代測定という異なる2つの方法を用いて決定した。 成長環のカウントには、暗色と明色の成長層を用いた2つの異なるプロトコルが使用された。 その結果、様々な結果が得られましたが、いくつかのサンゴでは、放射性炭素年代測定法とも矛盾していま した。 B. patula の放射性炭素年代は、この種のサンゴが長寿であることを示し、年代は 385 年以上、成長速度は線状(5.2-9.6mm/yr)、放射状(11.1-35.7μm/yr)と遅かったです。 これらの結果は、ニュージーランドの同地域や世界各地の他の黒サンゴ属の年齢や成長速度と比較可能である。 深海生物の年齢と成長率は、これらの生物が人為的攪乱に対する潜在的なリスクを判断するために重要である。 これらの結果は、今後のリスク評価やニュージーランドのサンゴ管理の枠組みにおける管理戦略に活用される。
Data Availability Statement
本研究で作成したすべてのデータセットは論文/補足資料に含まれている。
Author Contributions
研究の構想・設計にはPM、DT、HB、NHが貢献した。 PMは試料調製、切片画像解析、薄切片プロトコルの作成とゾーンカウント、放射性炭素試料の微粉砕を全て行った。 NHは放射性炭素試料の分析を行いました。 SFは放射性炭素の生データから年代を推定し、原稿を執筆した。 PMとDTは原稿の第一稿を執筆した。
Funding
This research was funded from Year Two of the Conservation Services Programme (CSP), Department of Conservation (DOC) Project POP2017-07 Objective to “Develop a methodology to determine the age and growth characteristics of key high-risk New Zealand deep-sea (cold-water) coral species.” (ニュージーランドの高リスクの深海サンゴ種の年齢と成長特性を決定する方法を開発する). 本誌の掲載料は、ニュージーランドNIWAより提供された。
利益相反
著者らは、本研究が利益相反の可能性と解釈される商業的または金銭的関係のない状態で行われたことを宣言する。
謝辞
この研究を支援してくださったニュージーランド自然保護省コンサベーション・サービス・プログラム・グループ、特にIan Angus、Kris Ramm、またFreydis Hjorvarsdottir(前CSP、現ニュージーランド第一産業省)に対して感謝します。 Helen Neil (NIWA) と Dan Sinclair (VUW)には、予備データの提供を支援していただいた。 NIWA の Sadie Mills と Diana Macpherson にはサンプル選定について、Owen Anderson には図 1 の作成について、Peter Horn には内部レビューについて、Rosemary Hurst には原稿について有益なコメントと助言をいただいたことに謝意を表する。 また、Jeremy Horowitz (James Cook University, Australia) にはサンプルの同定を、Dennis Opresko (Emeritus Smithsonian Institute) には黒サンゴの分類法改訂についてコメントをいただきました。 最後に、3名の査読者に感謝する。彼らの詳細かつ建設的な評価によって、本書は大いに恩恵を受けた。
Adkins, J. F., Griffin, S., Kashgarian, M., Cheng, H., Druffel, E. R. M., Boyle, E. A., and al. (2002). 深海珊瑚の放射性炭素年代測定。 Radiocarbon 44, 567-580. doi: 10.1017/s0033822200031921
CrossRef Full Text | Google Scholar
Althaus, F., Williams, A., Schlacher, T. A., Kloser, R. J., Green, M. A., Barker, B. A.、et al. (2009). 海山の深海珊瑚生態系に対する底引き網漁の影響は長期的である。 Mar. Ecol. Prog. Ser. 397, 279-294. doi: 10.3354/meps08248
CrossRef Full Text | Google Scholar
Alves, E. Q., Macario, K., Ascough, P., and Bronk Ramsey, C.(2018). “Impacts of deep coral ecosystem on the seamount is long-lasting”(海山における底曳網の影響)。 世界的な海洋放射性炭素リザーバー効果:定義、メカニズム、および展望。 Rev. Geophys. 56, 278-305. doi: 10.1002/2017RG000588
CrossRef Full Text | Google Scholar
Anderson, O., Tracey, D., Bostock, H., Williams, M., and Clark, M.(2014).,日本学術振興会特別研究員(PD)。 ニュージーランドEEZにおける保護対象サンゴ種の精緻な生息地適合性モデリング. 自然保護省のために作成されたNIWAクライアントレポート。 WLG2014-69No.46. Auckland: NIWA.
Google Scholar
Andrews, A. H., Cailliet, G. M., Kerr, L. A., Coale, K. H., Lundstrom, C., and DeVogelaere, A. P. (2005). “Investigations of age and growth for three deep-sea corals from the Davidson Seamount off central California,” in Cold-Water Corals and Ecosystems, eds A. Freiwald and J. M. Roberts (Berlin: Springer-Verlag), 1021-1038. doi: 10.1007/3-540-27673-4_51
CrossRef Full Text | Google Scholar
Andrews, A. H……. (2005年4月), Cordes, E. E., Mahoney, M. M., Munk, K., Coale, K. H., Cailliet, G. M., et al.(2002年)。 アラスカ湾に生息する深海性ゴルゴニアン(Primnoa resedaeformis)の年齢、成長、放射年代測定による検証。 Hydrobiologia 471, 101-110.
Google Scholar
Anthony, K. R. N., and Marshall, P. A. (2009)(2010). サンゴ礁と気候変動. オーストラリアの海洋気候変動の影響と適応に関するレポートカード2009. Gland: IUCN.
Google Scholar
Bronk Ramsey, C. (2001). 放射性炭素校正プログラムOxCalの開発。 Radiocarbon 43, 355-363. doi: 10.1017/s0033822200038212
CrossRef Full Text | Google Scholar
Cairns, S., Gershwin, L., Brook, F., Pugh, P., Dawson, E., Ocanþa, O, etc. (2009).・Cha. D. P. Gordon (New Zealand: Canterbury University Press), 59-101.
Google Scholar
Carreiro-Silva, M., Andrews, A. H., Braga-Henriques, A., de Matos, V., Porteiro, F. M. and Santos, R. S. (2013). アゾレス諸島の長寿黒サンゴLeiopathes sp.の成長速度のばらつき。 Mar. Ecol. Progr. Ser. 473, 189-199. doi: 10.3354/meps10052
CrossRef Full Text | Google Scholar
Chiswell, S. M., Bostock, H. C., Sutton, P. J. H., and Williams, M. J. M. (2015年). ニュージーランド周辺深海の物理海洋学:レビュー,New Zealand. J. Mar. Freshw. Res. 49, 1-32. doi: 10.1080/00288330.2014.992918
CrossRef Full Text | Google Scholar
Clark, M., Bowden, D., Baird, S., and Stewart, R. (2010). チャタムライズ北部の「墓場」複合海域の海山の底生生物多様性に対する漁業の影響. New Zeal. Aqu. Environ. Biodivers. 46:40.
Google Scholar
Clark, M., and Rowden, A. (2009). 深海トロールがチャタム海嶺の大型無脊椎動物群に与える影響(ニュージーランド). Deep Sea Res. I 56, 1540-1554. doi: 10.1016/j.dsr.2009.04.015
CrossRef Full Text | Google Scholar
Clark, M., Tracey, D., Anderson, O. and Parker, S.(2014). “Effects on deepwater trawling on macro-vertebrate assembrates on the seamount on Chatham Rise, New Zealand. 保護されているサンゴの生態リスクアセスメントの試行(Pilot Ecological Risk Assessment for Protected Corals). ウェリントン: National Institute of Water and Atmospheric Research, 32.
Google Scholar
Clark, M. R., Bowden, D. A., Rowden, A. A., and Stewart, R.(2019).[英語版のみ]. 15年後の海山における底引き網漁に対する底生生物群集の回復力を示す証拠はほとんどない。 Front. Mar. Sci. 6:63. doi: 10.3389/fmars.2019.00063
CrossRef Full Text | Google Scholar
Coleman, D. C. (1991). カーボンアイソトープテクニック. Cambridge, MA: Academic Press.
Google Scholar
Consalvey, M., MacKay, K., and Tracey, D. (2006). ニュージーランド地域の保護対象深海珊瑚種に関する情報レビュー。 NIWAクライアントレポート. WLG2006-85. 60. Auckland: NIWA.
Google Scholar
Druffel, E. R. M., Griffin, S., Witter, A., Nelson, E., Southon, J., Kashgarian, M., et al.(1995).深海珊瑚礁の情報レビュー。 Gerardia: Bristlecone Pine of the Deep-sea?(ジェラルディア:深海のブリストルコーン・マツ)? Geochim. Cosmochim. Doi: 10.1016/0016-7037(95)00373-8
CrossRef Full Text | Google Scholar
Druffel, E. R. M., King, L. L., Belastock, R. A., and Buesseller, K. O. (1990). 210Pbと他の同位体を用いた深海珊瑚の成長速度. Geochim. Cosmochim. Acta 54, 1493-1500.
Google Scholar
Fallon, S. J., Fifield, L. K., and Chappell, J. M. (2010). オーストラリア国立大学における放射性炭素年代測定の次の章:シングルステージAMSのステータスレポート。 Nuclear Instr. Methods Phys. Res. Sec. B 268, 898-901. doi: 10.1016/j.nimb.2009.10.059
CrossRef Full Text | Google Scholar
Freeman,D., and Cryer,M. (2019). “Current management measures and threats,” in The State of Knowledge of Deep-Sea Corals in the New Zealand Region, Chap.9, eds D. M. Tracey and F. Hjorvarsdottir (Auckland: NIWA Science and Technology Series).
Google Scholar<8171>
Gammon, M. J……………………, Tracey, D. M., Marriott, P. M., Cummings, V. J., and Davy, S. K. (2018). 深海サンゴSolenosmilia variabilisの海洋酸性化に対する生理的応答. PEERJ 6:e5236. doi: 10.7717/peerj.5236
PubMed Abstract | CrossRef Full Text | Google Scholar
Griffin, S., and Druffel, E. M. (1989)(2019). 深海珊瑚への炭素供給源. Radiocarbon 31, 533-543. doi: 10.1017/s0033822200012121
CrossRef Full Text | Google Scholar
Hitt, N. T., Sinclair, D. J., Fallon, S. J., Neil, H. L., Tracey, D. M., Komugabe-Dixson, A.、他 (2020). ニュージーランド産黒サンゴの成長と長寿。 Deep Sea Res. I 2020:103298. doi: 10.1016/j.dsr.2020.103298
CrossRef Full Text | Google Scholar
Komugabe, A. F., Fallon, S. J., Thresher, R. E., and Eggins, S. M. (2014). 深海黒サンゴから得られた現代のタスマン海表層貯水池年代. Deep-Sea Res. 99, 207-212. doi: 10.1016/j.dsr2.2013.05.024
CrossRef Full Text | Google Scholar
Komugabe-Dixson, A. F., Fallon, S. J., Eggins, S. M., and Thresher, R. E. (2016). 南西太平洋の循環における完新世中期の変化に関する放射性炭素の証拠。 Paleoceanography 31, 971-985. doi: 10.1002/2016PA002929
CrossRef Full Text | Google Scholar
Love, M.S., Yoklavich, M.M., Black, B.A., and Andrews, A.H. (2007). 黒サンゴ(Antipathes dendrochristos)コロニーの年齢と関連する無脊椎動物種に関するメモ。 Bull. Mar. 80, 391-400.
Google Scholar
Matsumoto, K. (2007). 放射性炭素を用いた世界海洋の循環年齢。 J. Geophys. 深海性ゴルゴニアンPrimnoa resedaeformisとParagorgia arboreaの形態と成長。 Mar. また、このような環境下での生物多様性の保全は、生物多様性保全の重要な課題である。 竹サンゴの成長速度の変化:完新世中期深海性ゴルゴニアン骨格の硬化年代学と放射性炭素年代測定(Keratosis sp: チャタム海嶺(ニュージーランド)産のオクトコラリア(Octocorallia)。 Facies 54, 151-166. doi: 10.1007/s10347-007-0129-x
CrossRef Full Text | Google Scholar
Pinkerton, M. H. (2016). ニュージーランドEEZにおける植物プランクトンのOcean Colour Satellite Observations, 1997-2016. 環境省. Wellington: NIWA.
Google Scholar
Prouty, N. G., Fisher, C. R., Demopoulous, A. W. J., and Druffel, E. R. M. (2016). ディープウォーター・ホライズン原油流出事故の影響を受けたディーシー・コーラルの成長速度と年齢。 Deep-Sea Res. II 129, 196-212. doi: 10.1016/j.dsr2.2014.10.021
CrossRef Full Text | Google Scholar
Prouty, N. G., Roark, E. B., Buster, N. A. and Ross, S. W. (2011). メキシコ湾における深海クロサンゴの成長速度と年齢分布. Mar. Ecol. Prog. Ser. 423, 101-115. doi: 10.3354/meps08953
CrossRef Full Text | Google Scholar
Reimer, P. J., Bard, E., Bayliss, A., Beck, J. W., Blackwell, P. G., Bronk Ramsey, C.他 (2013).深海における深海クロサンゴの成長速度と年齢分布 IntCal13とMarine13の放射性炭素年代較正曲線0-50,000年 cal BP. Radiocarbon 55, 1869-1887. doi: 10.2458/azu_js_rc.55.16947
PubMed Abstract | CrossRef Full Text | Google Scholar
Reimer, P. J., and Reimer, R.W. (2001). 海洋貯水池補正データベースとオンライン・インターフェース. Radiocarbon 43, 461-463. doi: 10.1017/s0033822200038339
CrossRef Full Text | Google Scholar
Roark, E. B., Guilderson, T. P., Dunbar, R. B., Fallon, S. J., and Mucclarone, D. A. (2009). タンパク質性深海珊瑚の極端な長寿化. Proc. Natl. Acad. Sci. U.S.A. 106, 5204-5208. doi: 10.1073/pnas.0810875106
PubMed Abstract | CrossRef Full Text | Google Scholar
Roark, E. B., Guilderson, T. P., Dunbar, R. B. and Ingram, B. L. (2006)(2010). 放射性炭素による年齢と成長率。 ハワイ産深海珊瑚礁。 Mar. Ecol. Progr. 327, 1-37.
Google Scholar
Sánchez, J., and Rowden, A. (2006). Sherwood, O. A., and Edinger, E. N. (2009). “Octocoral diversity on New Zealand Seamounts,” in Proceedings of the Presented at the 10th International Coral Reef Symposium, Okinawa.
Google Scholar
Sherwood, O. A., and Edinger, E. N. (2009). ニューファンドランド・ラブラドールの深海性ゴルゴニアン及びアンチパサリアンコーラルの年齢と成長率. Can. J. Fish. 66, 142-152. doi: 10.1139/f08-195
CrossRef Full Text | Google Scholar
Sherwood, O.A., Scott, D.B., Risk, M.J., and Guilderson, T.P. (2005). 深海性八腕類Primnoa resedaeformisにおける年輪の放射性炭素証拠。 Mar. Ecol. Prog. 301, 129-134. doi: 10.3354/meps301129
CrossRef Full Text | Google Scholar
Sikes, E., Burgess, S., Grandpre, R., and Guilderson, T. (2008). 深海珊瑚を用いたニュージーランド地域の現代深海年代の評価. Deep Sea Res. I 55, 38-49. doi: 10.1016/j.dsr.2007.10.004
CrossRef Full Text | Google Scholar
Stuiver, M.and Polach, H. A. (1977). 14Cデータの報告についての考察。 Radiocarbon 19, 355-363. doi: 10.1017/S0033822200003672
CrossRef Full Text | Google Scholar
Thresher, R., Rintoul, S. R., Koslow, J. A., Weidman, C., Adkins, J., and Proctor, C. (2004). オーストラリア南部における過去3世紀にわたる気候変動の海洋学的証拠。 Geophys. Res. Lett. 31:l07212.
Google Scholar
Thresher, R. E., MacRae, C. M., Wilson, N. C., and Gurney, R. (2007). 深海性ゴルゴニアン(Keratoisis spp.;Isididae)の骨格構成に及ぼす環境効果. Bull. Mar. Sci. 8, 409-422.
Google Scholar
Tracey, D., Bostock, H., and Shaffer, M. (2018)(2018). 保護された深海サンゴのエイジング法. エイジング研究のレビューと提言. DOC Contract 4527 GMC – Age and Growth of coral (POP2017-07). NIWAクライアントレポートWN2018035. Auckland: NIWA, 40.
Google Scholar
Tracey, D., Neil, H., Gordon, D., and O’Shea, S. (2003)(2003). 深海の年代記:ニュージーランド海域の深海珊瑚の加齢。 NIWA Water Atmosph. 11, 22-24.
Google Scholar
Tracey, D. M., and Hjorvarsdottir, F. (eds.) (2019). “The state of knowledge of deep-sea corals in the new zealand region,” in NIWA Science and Technology Series Number 84, (Auckland: NIWA),140.
Google Scholar
Tracey, D. M., Neil, H., Marriott, P., Andrews, A. H., Cailliet, G. M., and Sanchez, J. A. (2007). ニュージーランド海域における深海性タケノコサンゴ(イシダ科)2属の年齢と成長。 Bull. Mar. Sci. 81, 393-408.
Google Scholar
Williams, A., Schlacher, T. A., Rowden, A. A., Althaus, F., Clark, M. R., Bowden, D. A., et al. 海山の巨大底生生物群集はトロール漁の影響から回復していない。 Mar. Ecol. 31, 183-199. doi: 10.1111/j.1439-0485.2010.00385.x
CrossRef Full Text | Google Scholar
Williams, B., and Grottoli, A. G. (2010). また、δ15Nとδ13Cの安定同位体比の変化と古海洋の復元への示唆を示した。 Geochim. Cosmochim. Acta 74, 5280-5288. doi: 10.1016/j.gca.2010.06.026
CrossRef Full Text | Google Scholar
Williams, B., Risk, M. J., Ross, S. W., and Sulak, K. J. (2007). 深海の抗パタリアンからの安定同位体データ。 アメリカ南東部沿岸における 400 年間の記録. Bull. Mar. Sci. 81, 437-447.
Google Scholar
.
Leave a Reply