Frontiers in Marine Science
Introduzione
I coralli di acque profonde sono un gruppo molto vario di organismi marini e la regione della Nuova Zelanda è un hotspot per i coralli di acque profonde (Sánchez e Rowden, 2006; Cairns et al., 2009). Diversi studi hanno indagato gli effetti della pesca a strascico sugli assemblaggi di macroinvertebrati di profondità (compresi i coralli) nella regione del Pacifico meridionale (Althaus et al., 2009; Clark e Rowden, 2009; Clark et al., 2010, 2019; Williams et al., 2010; Anderson et al., 2014). Questi hanno dimostrato impatti apprezzabili della pesca a strascico su colonie e comunità di coralli d’altura a crescita lenta e fragile con poca o nessuna capacità di recupero. Gli impatti ambientali previsti dal cambiamento climatico, come il riscaldamento degli oceani e l’acidificazione degli oceani, dovrebbero anche minacciare la salute dei coralli di acque profonde con alcuni habitat che diventano troppo caldi per loro per prosperare in Anthony e Marshall (2009), o ridotti in pH e ioni carbonato, che probabilmente avranno un impatto sui coralli di acque profonde con scheletri a base di carbonato (Gammon et al., 2018). Per informare meglio le future valutazioni del rischio e la gestione delle risorse marine, una comprensione dell’età e della crescita dei coralli di acque profonde è fondamentale per determinare i tempi di rigenerazione dei coralli e la recuperabilità dopo le perturbazioni antropogeniche o altri danni naturali.
Una notevole quantità di lavoro è stata intrapresa nella regione della Nuova Zelanda sulla tassonomia dei coralli di acque profonde, e la distribuzione delle diverse specie e le loro preferenze ambientali, così come l’importanza del corallo di mare profondo per gli ecosistemi bentonici. Una revisione della nostra comprensione e conoscenza dei coralli di acque profonde nella regione della Nuova Zelanda è stata recentemente pubblicata (Tracey e Hjorvarsdottir, 2019). Questa revisione ha anche delineato le principali lacune nella nostra comprensione dei coralli di acque profonde, compresi i dati limitati sull’età e sulla crescita per molti generi e specie. In questo studio valutiamo l’età e il tasso di crescita del corallo nero protetto di acque profonde Antipatharian Bathypathes patula. L’età e il tasso di crescita di questo corallo non sono stati precedentemente studiati in questa regione. La selezione di questa specie si è basata sull’elenco di priorità della valutazione del rischio (Clark et al., 2014) e sulla revisione della letteratura (Tracey et al., 2018), unita alla disponibilità di campioni (posizione e numero totale), e ad altre ricerche complementari sul corallo nero in corso in Nuova Zelanda (Hitt et al., 2020).
Tracey et al. (2018) hanno delineato i metodi attuali che vengono utilizzati per invecchiare le specie di coralli di acque profonde. I vantaggi e gli svantaggi di ogni metodo sono stati discussi. I principali metodi che sono stati applicati con successo per misurare l’età e la crescita dei coralli di acque profonde sono (1) l’osservazione diretta, come le misurazioni in situ o gli esperimenti in acquario della crescita lineare o delle estensioni di superficie; il tasso di aggiunta dei polipi; la stima dei tassi di calcificazione (ad es, utilizzando la tecnica del peso galleggiante), (2) enumerazione delle zone di crescita scheletrica o delle bande (sclero-cronologia), e (3) analisi radiometriche.
Le osservazioni dirette e le misurazioni in situ o in acquario sono molto difficili da intraprendere per i coralli di acque profonde, poiché è difficile ottenere esemplari vitali vivi e mantenerli in vita, poiché richiedono molto tempo per crescere (Gammon et al., 2018; Tracey et al., 2018). L’enumerazione delle zone di crescita, Sclerochronology, è ideale per le specie di coralli di acque profonde che hanno un alto contrasto tra i modelli di crescita a zone osservati nelle sezioni trasversali scheletriche. È essenziale, tuttavia, che un metodo di convalida indipendente sia usato per testare la validità cronologica dei conteggi delle zone di crescita osservate (Andrews et al., 2002). Il metodo per ottenere il conteggio delle zone di crescita visibili nella struttura scheletrica è stato applicato con successo per determinare i dati sull’età e il tasso di crescita di octocoralli gorgonie tra cui coralli di bambù Keratoisis sp. di 400 anni, Primnoa spp. di 100 anni, e Lepidisis sp. di 60 anni (Thresher et al., 2004, 2007; Mortensen e Buhl-Mortensen, 2005; Sherwood et al., 2005; Tracey et al., 2007; Sherwood e Edinger, 2009). I coralli neri sono stati anche invecchiati con successo usando la zona di crescita o i conteggi, con età da 150 anni a 480 anni per Antipathes dendrochristos e Leiopathes glaberrima, rispettivamente (Love et al., 2007; Williams et al., 2007). In uno studio sul corallo nero Stauropathes arctica, sono stati ottenuti conteggi di zona di 55-58 anni (Sherwood e Edinger, 2009), con gli autori che notano che la datazione al radiocarbonio ha limitato queste età a 55 e 82 anni. I tassi di crescita dei coralli neri tramite l’enumerazione delle zone di crescita rivelano una bassa crescita radiale da 8 a 140 μm/anno (Love et al., 2007; Prouty et al., 2011).
Ci sono potenziali limitazioni al metodo di enumerazione delle zone di crescita (vedi sintesi in Tracey et al., 2018). Molti coralli non mostrano una chiara struttura a zone regolari in tutto il loro scheletro, o la chiara struttura a zone può essere evidente solo in porzioni dello scheletro. Anche quando è presente, la struttura a zone può essere variabile o mostrare diverse possibilità di interpretazione. Per esempio, i conteggi basali e di punta delle specie di ottocoralli di bambù Keratoisis sp. e Lepidisis sp. erano superiori alle età stimate con il metodo radiometrico del piombo 210 (210Pb) (Tracey et al., 2007). Si è ipotizzato che le zone osservate al microscopio ottico abbiano una periodicità biennale e che le zone osservate al SEM alla giunzione nodale possano rappresentare un evento ambientale, come la periodicità lunare. I cicli lunari possono essere associati a forti eventi di marea, che potrebbero aumentare la disponibilità di cibo per gli organismi sessili che si nutrono di filtri, dando luogo a impulsi di crescita somatica. I coralli bubblegum gorgoniani (ad esempio, Paragorgia spp.) e i coralli rossi preziosi (ad esempio, Corallium sp.) non sono stati invecchiati con successo usando i conteggi di zona a causa di modelli di bande intrinsecamente complessi o poco definiti (Griffin e Druffel, 1989; Andrews et al, 2005).
Il metodo radiometrico più comune utilizzato nei coralli di acque profonde è la datazione con l’isotopo del radiocarbonio (14C) (Druffel et al., 1995; Sherwood et al., 2005; Tracey et al., 2018). Questo metodo si basa sul fatto che una piccolissima quantità di carbonio naturale nell’atmosfera è radioattiva sotto forma di 14C e viene incorporata nello scheletro del corallo quando forma il suo carbonato di calcio o proteine e chitina (Adkins et al., 2002; Tracey et al., 2003; Consalvey et al., 2006). Poiché l’emivita del radiocarbonio è nota per essere di 5.730 anni, questo metodo, anche se costoso, può essere utilizzato per invecchiare in modo affidabile campioni di meno di 50.000 anni fa; oltre questa data, l’attività del 14C diventa troppo bassa per essere rilevata (Coleman, 1991). Durante gli anni 1950-60, c’è stato un rapido aumento del 14C atmosferico derivante dai test di dispositivi nucleari; questo “carbonio bomba” può anche essere usato come strumento per calibrare le età, fornendo un punto di riferimento per campioni più recenti (Coleman, 1991; Druffel et al., 1995; Tracey et al., 2003; Roark et al, 2009; Sherwood e Edinger, 2009).
La datazione al radiocarbonio è stata utilizzata con successo su diverse specie di coralli, tra cui coralli duri, coralli neri e octocoralli gorgonie (Druffel et al., 1990; Roark et al., 2006; Carreiro-Silva et al., 2013; Prouty et al., 2016; Hitt et al., 2020). Tuttavia, ci sono anche delle limitazioni con questo metodo. Come risultato della circolazione oceanica, l’età radiocarbonica dell’acqua negli oceani è significativamente più vecchia di quella dell’atmosfera, la cosiddetta “età della riserva marina” (Tracey et al., 2003; Consalvey et al., 2006; Roark et al., 2006; Noé et al., 2008). Errori nell’età del serbatoio possono avere effetti significativi sulle stime dell’età dei coralli. La correzione delle età del serbatoio richiede la conoscenza delle acque circostanti e presuppone rapporti costanti di radiocarbonio nell’acqua; tuttavia, questi livelli possono essere alterati dalla miscelazione all’interno dell’oceano, da eventi di upwelling localizzati e da fioriture di produttività, tutti elementi che possono alterare le trasformazioni del ciclo del carbonio e la composizione scheletrica dei coralli del 14C (Tracey et al., 2018).
In base alla revisione di Tracey et al. (2018) abbiamo scelto di utilizzare due metodi di invecchiamento per B. patula; i conteggi delle bande di crescita scheletrica e il metodo radiometrico di datazione al radiocarbonio (14C). L’applicazione di questi metodi permette il confronto con altri studi precedenti sull’invecchiamento dei coralli sia in Nuova Zelanda che a livello globale. Vengono presentati la selezione dei campioni, la microfresatura del materiale e i metodi di sezionamento, nonché l’interpretazione del conteggio delle zone e della datazione al radiocarbonio. Vengono fornite le caratteristiche di età e crescita del corallo nero B. patula e di una colonia di Bathypathes cf. conferta (inizialmente assunta come B. patula).
Materiali e metodi
Selezione dei campioni
I campioni utilizzati in questo studio provengono da esemplari conservati presso il National Institute of Water and Atmospheric Research (NIWA) della Nuova Zelanda, National Invertebrate Collection (NIC).
I campioni sono stati selezionati da due regioni; la Bay of Plenty (nord-est dell’Isola del Nord della Nuova Zelanda) e Chatham Rise (est della Nuova Zelanda), per confrontare i risultati e valutare la variabilità dei tassi di crescita tra le diverse aree, e anche con altri generi di corallo nero di profondità, in particolare Antipathella, Leiopathes, e Antipathes, dalle stesse regioni (Hitt et al., 2020).
Le correnti oceanografiche influenzano i coralli di profondità fornendo un flusso costante di cibo e la dispersione delle larve. Di conseguenza, queste correnti influenzano anche la disponibilità di prede e la distribuzione dei coralli di acque profonde. Mentre i coralli di profondità sono più influenzati dalle correnti di acqua di fondo che scorrono su di loro e influenzano la distribuzione dei sedimenti, sono anche influenzati dalle correnti di acqua superficiale che influenzano la produttività primaria e quindi la fornitura di cibo che piove fino al corallo di profondità sul fondo del mare. La massa della Nuova Zelanda si trova all’incrocio tra le acque superficiali subtropicali, calde e povere di nutrienti, che scorrono da nord, e le acque superficiali subantartiche, fresche e ricche di nutrienti, che scorrono da sud. L’attuale comprensione delle masse d’acqua e delle correnti oceaniche della regione in superficie, del termoclino, delle acque intermedie e delle acque profonde, è stata recentemente rivista da Chiswell et al. (2015). L’impostazione oceanografica per i coralli da questo studio sono i seguenti.
La Baia di Plenty è bagnata da acque superficiali calde, a basso contenuto di nutrienti, subtropicali che provengono dalla corrente orientale australiana e fluiscono da ovest e formano la corrente orientale di Auckland. Il Chatham Rise è il punto d’incontro tra le acque subtropicali (STW), calde e povere di nutrienti, e le acque subantartiche (SAW), fresche e ricche di nutrienti, provenienti dall’Oceano Meridionale, il fronte subtropicale (STF). La miscelazione delle masse d’acqua STW e SAW al STF fa sì che il Chatham Rise sia una zona di altissima produttività primaria che fornisce una significativa fonte di cibo e quindi di carbonio per questi coralli, le concentrazioni di clorofilla a variano da 0,3 e 0,48 mg/m3 nel Chatham Rise e Bay of Plenty, rispetto a 0,03-0,3 mg/m3 nel Pacifico centrale (Pinkerton, 2016). A profondità di 500-1500 m le acque intermedie antartiche (AAIW) fresche (3,5-10°C), a bassa salinità (34,4-34,6) e ad alto contenuto di ossigeno (200-250 μmol/kg) e di nutrienti (nitrati 15-36 μmol/kg) fluiscono in questa regione da nord e da sud (Chiswell et al, 2015).
Sono stati scelti i coralli del Chatham Rise perché ci sono dati moderni di età del serbatoio 14C da questa zona (Sikes et al., 2008). Conoscere l’età della riserva locale di 14C permette una stima più precisa dell’età radiocarbonica dei coralli.
I campioni sono stati selezionati da esemplari esistenti raccolti dalla Fisheries New Zealand (FNZ), da osservatori del Ministry for Primary Industries (MPI) e da ricercatori NIWA e sono conservati presso il (NIC). Un grafico dei dati di distribuzione di B. patula ha aiutato la decisione sulla selezione del sito della colonia e il numero di campioni (Figura 1). Due esemplari della Baia di Plenty (NIWA24190 e NIWA85940) sono stati selezionati per la datazione al radiocarbonio e le sezioni sottili per il conteggio delle zone. Il resto degli esemplari sono stati selezionati dal Chatham Rise.
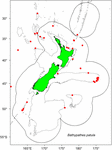
Figura 1. Località in cui Bathypathes patula è stata campionata nella regione della Nuova Zelanda. I punti rossi indicano i luoghi di provenienza dei campioni di corallo per gli esemplari di B. patula conservati nella National Invertebrate Collection of New Zealand (NIC). Le linee grigie denotano i 200 m di profondità dell’acqua. La linea nera denota l’estensione della zona economica esclusiva della Nuova Zelanda (EEZ).
A fine 2018, uno specialista in visita nella tassonomia del corallo nero (Jeremy Horowitz, James Cook University) ha rivisto le identificazioni di diversi campioni nel NIC. Durante questo processo una delle colonie di corallo nero selezionate per questa ricerca di stima dell’età e assunta come Bathypathes patula, ha ricevuto un’identificazione rivista. Il corallo nero NIWA24190 è stato ora identificato come Bathypathes cf. conferta. Il campione è stato indicato come Bathypathes cf. conferta in tutto questo documento (Figura 2), l’identificazione più probabilmente corretta. Un’ulteriore revisione tassonomica di questo campione è in corso (Opresko pers comm). La mappa di distribuzione (Figura 1) può includere ulteriori esemplari di B. cf. conferta.

Figura 2. Bathypathes cf. conferta. NIWA24190. Campione che mostra dove sono stati estratti i campioni per l’analisi. (A) è la punta di crescita, (B) la sezione basale. Esemplare proveniente da Bay of Plenty, Nuova Zelanda. La barra della scala è un righello di 35 cm.
Preparazione del materiale
Una volta identificati i campioni adatti, sono state estratte piccole sezioni (meno di 10 mm) dalle punte di crescita e dalla parte basale del gambo principale dei coralli. Poiché molti dei campioni erano stati rotti durante il processo di raccolta, sono state prese ulteriori sezioni lungo il gambo principale in modo da poter generare stime affidabili del tasso di crescita lineare (Figure 2-5). I campioni dalle punte di crescita sono stati poi divisi in due frammenti, uno per la datazione radiocarbonica e l’altro per la preparazione di sezioni sottili per ottenere il conteggio delle zone.

Figura 3. Bathypathes patula NIWA85940. Campione che mostra dove sono stati estratti i campioni per l’analisi. (A) è la punta di crescita, (C) la sezione basale, e (B) è un sito di campionamento intermedio poiché l’esemplare era rotto al momento della raccolta e lunghezze sconosciute del gambo principale possono mancare sopra questo sito. Esemplare da Bay of Plenty, Nuova Zelanda. La barra della scala è un righello di 35 cm.

Figura 4. Bathypathes patula NIWA49468. Campione che mostra dove sono stati estratti i campioni per l’analisi. (A) è la punta di crescita, (D) la sezione basale, e (B,C) sono siti di campionamento intermedi in quanto l’esemplare era rotto al momento della raccolta e una lunghezza sconosciuta del ramo principale può mancare tra questi due siti. Esemplare da Chatham Rise, Nuova Zelanda. La barra della scala è un righello di 35 cm.

Figura 5. Bathypathes patula NIWA47911. Campione che mostra dove sono stati estratti i campioni per l’analisi. (A) è la punta di crescita, (F) la sezione basale e, (B-E) sono siti di campioni intermedi presi per cercare il segnale della bomba radiocarbonica. Esemplare proveniente da Chatham Rise, Nuova Zelanda. La barra della scala è un righello da 35 cm.
Micro-fresatura del materiale
Dal lavoro precedente è stato notato che le sezioni di corallo nero sono inclini a disegnare la resina attraverso la loro matrice porosa per azione capillare e quindi le sezioni in questo studio non potevano essere legate con la resina a una piastra di base per la fresatura. È stato invece realizzato un mandrino per tenere la sezione durante il processo di fresatura, eliminando così il rischio che la resina contaminasse i campioni di radiocarbonio.
I campioni di radiocarbonio in polvere sono stati estratti dal bordo e dal nucleo delle sezioni basali, centrali e di punta utilizzando una micromill New WaveTM con una fresa Brassler H2.11.006 da 0,5 mm presso NIWA. I campioni del bordo comprendevano materiale dai 250 micron esterni della sezione, i campioni del nucleo sono stati ottenuti da materiale entro un raggio di 750 micron dal primordio. Abbiamo cercato di estrarre almeno 1,5 mg di materiale per campione. I campioni sono stati poi pesati e analizzati per il radiocarbonio presso il laboratorio di radiocarbonio dell’Australian National University (ANU).
Metodo di sezionamento e protocolli di conteggio delle zone
Le sezioni del gambo del corallo sono state incorporate in resina epossidica trasparente in due parti e sezionate con una sega a sega diamantata alla NIWA. Le sezioni sono state lucidate su un lato e montate con il lato lucidato verso il basso su un vetrino per microscopia, quindi smerigliate e lucidate fino ad ottenere uno spessore ottimale per la visualizzazione della struttura della zona di crescita utilizzando la luce trasmessa. Lo spessore ottimale per le sezioni di questa specie era di circa 250-300 μm e segue la procedura standard per la preparazione delle sezioni sottili (Andrews et al., 2002; Mortensen e Buhl-Mortensen, 2005; Tracey et al., 2007). Il conteggio delle bande di crescita periodica o degli incrementi, che si presume si formino annualmente, ha avuto luogo dopo la preparazione della sezione sottile al NIWA.
Per generare i conteggi delle zone, le preparazioni delle sezioni dei rami principali sono state viste al microscopio composto con luce trasmessa. Come aiuto all’interpretazione delle zone, le sezioni sono state viste anche con luce ultravioletta seguendo la metodologia di Sherwood e Edinger (2009). In questo caso i conteggi delle zone sono stati fatti utilizzando la luce trasmessa in campo chiaro e l’illuminazione ultravioletta è stata un utile aiuto per aiutare a definire ulteriormente la struttura delle zone osservate.
Metodo di datazione al radiocarbonio
Prima delle analisi, i campioni sono stati puliti (ad esempio, lisciviati con acido) dalle croste nere e dall’attività endolitica per rimuovere qualsiasi potenziale contaminante più giovane 14C che potrebbe alterare i risultati (Adkins et al., 2002). I campioni di corallo in polvere sono stati trattati con <1 ml di 0,1 M HCL. L’acido è stato poi rimosso con una pipetta e i campioni sono stati lavati tre volte con acqua milliQ. Il campione è stato poi congelato con ghiaccio secco e liofilizzato. Circa 1 mg di campione di corallo pulito è stato pesato in una provetta di quarzo da 6 mm OD, sono stati aggiunti ∼60 mg di CuO e una coppa d’argento. I tubi sono stati evacuati a <3e-3 torr (<0,4 Pa) e sigillati con una torcia. I tubi sono stati poi cotti a 900°C per 6 ore per generare CO2. La CO2 risultante è stata purificata e convertita in grafite in presenza di idrogeno con Fe alimentato come catalizzatore. La grafite è stata poi misurata sullo spettrometro di massa con acceleratore a stadio singolo dell’ANU (Fallon et al., 2010). Tutti i dati sono stati corretti utilizzando on-line AMS δ13C, normalizzato per l’acido ossalico I e sfondo sottratto utilizzando 14C carbone libero trattato nello stesso modo come i campioni di corallo. I dati sono presentati secondo le raccomandazioni di Stuiver e Polach (1977).
Risultati
Sezioni sottili per i conteggi di zona
I modelli strutturali di zona osservati sulle sezioni scheletriche per queste specie sono molto complessi e possono essere interpretati in molti modi. Per il conteggio della crescita radiale le sezioni preparate sono state osservate attentamente e l’asse che meglio mostrava una chiara struttura a zone dal primordio al bordo esterno è stato poi utilizzato per ottenere il conteggio delle zone. La crescita radiale dei rami in B. patula era ragionevolmente regolare, non c’era una marcata distorsione nella deposizione delle zone che formavano zone più spesse su un lato particolare del ramo, che è stato precedentemente osservato in altre specie di corallo nero. I raggi utilizzati per i calcoli del tasso di crescita erano una media dei raggi minimi e massimi osservati nelle sezioni preparate, il che significa che i tassi di crescita radiali riportati saranno un tasso di crescita radiale medio per il corallo.
Sullo scheletro del corallo c’è un modello iniziale a bande grossolane in gran parte definito da un’alternanza di zone più scure e più chiare (Figura 6) quando si osserva sotto bassa potenza (10-20×). Ad un esame più attento sotto una potenza superiore (100-200×), c’è anche un modello di bande fini ragionevolmente regolare apparente (Figura 7). Qui abbiamo applicato due protocolli di lettura – contando sia le zone larghe grossolanamente formate sia le zone sottili finemente formate (Tabella 1). I conteggi delle zone qui presentati sono stati fatti da un solo lettore (Marriott). Durante il processo di sviluppo dei protocolli di interpretazione delle zone, sono state effettuate molte letture applicando le varie iterazioni dei protocolli in fase di sviluppo. I conteggi delle zone dei due protocolli qui presentati sono stati derivati da conteggi multipli in tutta la sezione.

Figura 6. Sezione basale del corallo NIWA47911 che mostra l’interpretazione delle bande colorate grossolane della struttura a zone osservata. L’esemplare mostra 54 zone contrassegnate da punti rossi. Il rettangolo tratteggiato nell’inserto mostra la porzione centrale della sezione basale ripresa nella Figura 7.

Figura 7. Porzione centrale della sezione basale del corallo NIWA47911 che mostra il protocollo di interpretazione a scala fine della struttura a zone osservata. L’esemplare mostra 42 zone marcate con punti neri in questa regione interna della sezione; l’intera sezione mostra 211 zone.

Tabella 1. Conteggio delle zone dalle sezioni sottili.
Per la parte più giovane dei campioni di studio, la punta o la regione vicina alla punta, il conteggio variava da 4 a 7 (conteggio delle zone grossolane) e da 10 a 19 (conteggio delle zone fini). Nella regione centrale del ramo principale, i conteggi variavano da 55 a 60 (conteggi della zona grossolana) e da 370 a 374 (conteggi della zona fine). Per la regione della base della colonia i conteggi variavano da 37 a 193 (conteggi delle zone grossolane) e da 78 a 481 (conteggi delle zone fini). La complessità dei modelli di zonazione ha evidenziato la necessità di una convalida.
Analisi al radiocarbonio
I campioni macinati elencati nella Tabella 2 sono stati inviati all’ANU per la datazione al radiocarbonio. I risultati dei 17 campioni di corallo sono riportati nei risultati del radiocarbonio (Tabella 3). Tre campioni (i campioni 47911-F2; 24190-B1; 85940-B1) non sono stati esaminati in laboratorio e non è rimasto materiale sufficiente per tentare un duplicato. I risultati del radiocarbonio dei coralli rimanenti sono stati calibrati su un’età solare utilizzando OxCal4.3 (Bronk Ramsey, 2001), la curva Marine13 e un deltaR locale neozelandese (offset del serbatoio del radiocarbonio) di -18 ± 36 (Reimer e Reimer, 2001). Le distribuzioni degli intervalli di età per i campioni di corallo pre-bomba sono mostrate nella Tabella 3. Tutti gli esemplari di corallo utilizzati in questo studio erano vivi al momento della cattura, quindi le stime dell’età fornite si riferiscono all’età al momento della cattura e non alla durata della vita dell’esemplare. La durata della vita prevista per questa specie sarà superiore alle stime di età qui presentate. Il corallo 85940 aveva circa 380 ± 50 anni, il corallo 49468 circa 310 ± 70 anni e il corallo 47911 circa 120 ± 50 anni (Tabella 3). Tutti i campioni del bordo esterno e della punta dei coralli contenevano valori di radiocarbonio F14C >1 (Tabella 3), ad eccezione di 85940-B2 e 85940-C2, il che indica che questi coralli erano vivi e incorporavano il carbonio dell’acqua superficiale al momento della raccolta. I coralli 85940-B2 e 85940-C2 avevano valori di radiocarbonio del bordo esterno F14C <1 (Tabella 3) che indicano che una parte dello scheletro non stava aggiungendo attivamente materiale. Questo è stato osservato in precedenza dove la parte basale della colonia corallina smette di crescere, tuttavia la sezione superiore continua a crescere (Komugabe-Dixson et al., 2016).
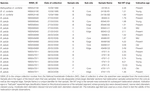
Tabella 2. Campioni estratti per la datazione al radiocarbonio.

Tabella 3. Risultati del radiocarbonio.
Stime del tasso di crescita
Le stime del tasso di crescita si sono basate sui risultati del radiocarbonio, poiché il conteggio delle zone si è rivelato problematico e non validato, quindi non poteva essere utilizzato per generare stime affidabili del tasso di crescita (vedere la sezione “Stime di età e crescita di Bathypathes patula” per ulteriori discussioni sul conteggio delle zone). I tassi di crescita sono stati derivati dalle distanze fisiche tra i punti di campionamento e dalle età radiocarboniche calibrate nel relativo sito di campionamento. A causa del fallimento di alcuni campioni durante l’analisi radiocarbonica, è stato possibile generare solo una serie limitata di stime dei tassi di crescita. I tassi di crescita radiale, il tasso al quale il ramo della colonia si ispessisce radialmente nel tempo, sono stati stimati dividendo gli anni solari calibrati di crescita tra il nucleo e il bordo del sito del campione per il raggio medio del segmento del ramo. La crescita radiale variava da 11,1 a 35,7 μm/anno. I tassi di crescita lineare, il tasso al quale il ramo della colonia si allunga longitudinalmente nel tempo, sono stati stimati dividendo la differenza tra le età di calendario calibrate dei campioni del nucleo per la distanza lineare tra due siti di campionamento. Sono stati ottenuti due tassi di crescita lineare, 5,2 e 9,6 mm/anno (Tabella 4).

Tabella 4. Stime del tasso di crescita di Bathypathes patula.
Discussione
Stime di età e crescita di Bathypathes patula
I risultati del radiocarbonio di questo lavoro mostrano che B. patula è una specie longeva, con età superiori a 385 anni. I tassi di crescita lineare (5,2-9,6 mm/anno) e radiale (11,1-35,7 μm/anno) sono stati ottenuti dai dati sull’età radiocarbonica.
Nessuno dei due protocolli di conteggio delle zone si è correlato in modo affidabile alle stime indipendenti dell’età radiocarbonica. Il protocollo di conteggio delle zone a scala fine ha mostrato la correlazione più stretta con i dati dell’età 14C, fornendo un supporto generale alla longevità quando si confrontano i conteggi delle zone con le date 14C. Le stime dell’età tra i dati del radiocarbonio e i dati del conteggio delle zone a scala fine hanno mostrato regolarmente una disparità di più di cento anni nelle sezioni dei rami con età radiocarboniche calibrate di 112-385 anni (Tabelle 1, 3). Le differenze nelle età stimate da campioni comparabili utilizzando i due metodi sono state negative e positive; non c’è stata alcuna sotto- o sovra-età utilizzando i conteggi di zona in misura proporzionale (Tabella 1). Questo significa che la struttura a zone osservata non mostra una periodicità annuale, o qualsiasi altro modello cronologico regolare, e i nostri protocolli di conteggio a zone non possono essere utilizzati per generare stime affidabili dell’età di B. patula.
I risultati del radiocarbonio sul bordo del campione NIWA-85940 suggeriscono che la sezione basale (C2) del fusto principale ha smesso di depositare materiale scheletrico sulla sua superficie esterna 293 anni prima della raccolta, e la sezione centrale (B2) ha smesso di depositare materiale scheletrico 88 anni prima della raccolta. Questo nonostante l’osservazione che queste sezioni di gambo del ramo erano immediatamente adiacenti a pinnule laterali con polipi vivi. L’aspetto ottico di queste sezioni non dava alcuna indicazione che la deposizione di zone fosse cessata sul loro margine esterno. La superficie del ramo del corallo sembrava essere ancora relativamente incontaminata, non mostrava alcun segno di erosione superficiale o scolorimento, come ci si potrebbe aspettare in un ramo di corallo che non stava più accrescendo nuovo materiale scheletrico sulla sua superficie esterna. Questo suggerisce che se i gambi dei rami possono smettere di accrescere gli strati scheletrici, mentre le estremità dei rami continuano a crescere, allora i protocolli del metodo di conteggio a zone non possono essere utilizzati in modo affidabile per la generazione di stime di età dalle sezioni basali per questa specie di corallo.
Nel campione NIWA47911 cinque campioni sono stati estratti dalla punta crescente del ramo assiale principale verso la base del corallo. Questi campioni sono stati distanziati di 25 mm. Questo era un tentativo di localizzare la comparsa del radiocarbonio bomba nello scheletro del corallo. In questo modo, il tasso di crescita lineare poteva essere valutato con precisione per le punte di crescita più recenti dei rami della colonia. Se l’aumento del radiocarbonio della bomba potesse essere collocato con precisione, allora fornirebbe anche un punto di riferimento preciso per impostare la correzione del 14C da applicare ai campioni di radiocarbonio pre-bomba per questa colonia e per altre colonie della stessa massa d’acqua. Sfortunatamente, tutti questi campioni si sono rivelati post-bomba, quindi questo processo non ha potuto essere completato. Un futuro campionamento più indietro lungo questo ramo sarebbe utile per stabilire questo punto di legame e il tasso di crescita lineare contemporaneo. Il nostro lavoro ha misurato un tasso di crescita lineare complessivo di 5,2-9,6 mm/anno, ipotizzando il tasso di crescita più veloce, sarebbe necessario campionare 700 mm lungo il ramo principale dalla punta di crescita per essere sicuri di osservare l’aumento del radiocarbonio della bomba nello scheletro.
Anche la ricerca di Love et al. (2007) sulla specie di corallo nero Antipathes dendrochristos ha riportato due interpretazioni della struttura della zona simili ai protocolli grossolano e fine qui descritti. Tuttavia, questi autori hanno scoperto che la struttura a bande colorate più grossolane era correlata ai loro risultati del radiocarbonio e del 210Pb che indicavano bande annuali (Love et al., 2007).
Le misurazioni del radiocarbonio del presente lavoro hanno mostrato F14C >1 nei campioni provenienti dalle punte dei rami in crescita e dalla superficie del ramo principale, indicando che questi campioni mostrano livelli moderni di radiocarbonio post-bomba. La presenza di carbonio post-bomba al di sotto dello strato misto dell’oceano, indica che i coralli neri B. patula stanno utilizzando POC di acqua superficiale esportato di recente. A queste profondità il POC è l’unica fonte di carbonio marcato con il 14C della bomba. Il carbonio inorganico disciolto (DIC) dell’acqua ambiente in cui vivono i coralli ha valori di 14C molto più bassi (Matsumoto, 2007; Alves et al., 2018). Rilevare il 14C bomba nei coralli che crescono in acque che non hanno sperimentato il 14C bomba (cioè, attraverso la ventilazione oceanica) rafforza che i coralli neri derivano il loro carbonio dal POC derivato dalla superficie (Roark et al., 2009; Sherwood e Edinger, 2009; Williams e Grottoli, 2010; Prouty et al., 2011). Questo significa anche che non abbiamo bisogno di modellare per un effetto serbatoio in acque profonde, come nel caso dei coralli duri, che derivano il loro carbonio scheletrico dal DIC nelle acque intermedie che li bagnano.
Confronti con i recenti dati neozelandesi e globali sull’età del corallo nero
Le stime dell’età derivate dal radiocarbonio e i tassi di crescita di questo progetto sono simili ai risultati di una serie di altri studi sull’età e la crescita che sono stati recentemente intrapresi per i coralli neri di profondità (Antipatharia) nella regione della Nuova Zelanda (Tabella 5). Queste stime di età e crescita per B. patula sono simili ad altri generi di corallo, come le colonie di Leiopathes, Antipathes e Antipathella, che vivono fino a 2600 anni e crescono dell’ordine di 1-143 μm/anno (Hitt et al., 2020). I nostri risultati forniscono un ulteriore supporto al fatto che i coralli neri neozelandesi sono alcuni degli animali marini a crescita più lenta della regione, con alcune specie che mostrano una longevità estrema.

Tabella 5. I risultati di B. patula e di altri coralli neri invecchiati in Nuova Zelanda non mostrano chiare tendenze nei tassi di crescita o nella longevità rispetto alle masse d’acqua e alle profondità in cui questi coralli risiedono. I campioni provenienti da Chatham Rise e Bay of Plenty provengono per lo più da profondità d’acqua ragionevolmente simili, tra 750 e <1269 m. L’esemplare più profondo di B. patula di Chatham Rise proviene da qualche parte tra 1120 e 1269 m (Tabelle 1, 5). Il tasso di crescita radiale dei nostri campioni di Chatham Rise (NIWA-49468 e NIWA47911) a 20-36 μm/anno è stato superiore a quello del campione di Bay of Plenty (NIWA-85940) a 11 μm/anno. Poiché solo tre tassi di crescita sono stati ottenuti con successo per i campioni di B. patula da queste due regioni, non si possono fare ipotesi sulle differenze regionali nei tassi di crescita. Lo spettro dei dati sul tasso di crescita per i coralli neri di Chatham Rise mostra che un campione di Antipathes sp. ha avuto il più veloce tasso di crescita osservato finora, 91-143 μm/anno, così come uno dei più lenti, 5-15 μm/anno per un campione di Leiopathes secunda. Bay of Plenty esemplari di corallo nero Bathypathes patula, Leiopathes secunda, e Leiopathes sp. mostrano medio a basso tasso di crescita radiale, 5-40 μm / anno. Al contrario, il corallo nero delle Fiordland in acque poco profonde Antipathella fiordensis ha un tasso di crescita radiale relativamente alto (50-57 μm/anno), ma non è chiaro se questa sia una funzione della specie o delle condizioni ambientali del fiordo, o una miscela di entrambi.
Questa ricerca concorda anche con gli studi globali che hanno dimostrato che i coralli neri sono alcuni dei coralli di mare profondo a crescita più lenta. Sherwood e Edinger (2009) danno stime di età per il corallo nero Stauropathes arctica fino a 82 anni solari. Prouty et al. (2011) hanno usato il radiocarbonio per generare stime dell’età delle colonie di Leiopathes sp. vive alla cattura fino a 2100 anni. Carreiro-Silva et al. (2013) hanno invecchiato una colonia di Leiopathes sp. delle Azzorre a 2320 anni. Un esemplare di Leiopathes annosahas delle Hawaii ha mostrato il potenziale di raggiungere una durata di vita superiore a 4000 anni (Roark et al., 2009).
Questi nuovi tassi di età e di crescita per il delicato corallo nero di acque profonde B. patula evidenziano che si tratta di una specie longeva, dimostrata in questo studio per raggiungere età superiori a 385 anni, e mostra lenti tassi di crescita lineare e radiale. I risultati indicano una probabile bassa resilienza e un lento tempo di recupero dopo un disturbo/rimozione. Questa ricerca sarà utilizzata per informare le future valutazioni del rischio e le strategie di gestione nel quadro della Nuova Zelanda per la gestione dei coralli (Freeman e Cryer, 2019). La crescita lenta e l’estrema longevità evidenzia ulteriormente che questi coralli saranno altamente vulnerabili alle perturbazioni sottomarine, come la pesca e l’estrazione mineraria, e potrebbero richiedere secoli per recuperare dagli impatti antropogenici (Hitt et al, 2020).
Conclusione
L’età e il tasso di crescita di campioni di corallo nero di acque profonde B. patula provenienti da due regioni della Nuova Zelanda, Bay of Plenty e Chatham Rise, sono stati determinati utilizzando due metodi diversi: conteggio degli anelli di crescita (sclerocronologia) e datazione al radiocarbonio. Sono stati utilizzati due diversi protocolli per contare gli anelli di crescita, utilizzando strati di crescita scuri e chiari. Questi hanno fornito risultati diversi, che erano anche incoerenti con le età radiocarboniche su molti di questi coralli. Le età al radiocarbonio di B. patula indicano che questa specie di corallo vive a lungo, con età di oltre 385 anni, con un lento tasso di crescita lineare (5,2-9,6 mm/anno) e radiale (11,1-35,7 μm/anno). Questi risultati sono comparabili con le età e i tassi di crescita di altri generi di corallo nero delle stesse regioni della Nuova Zelanda e a livello globale. L’età e i tassi di crescita delle specie di acque profonde sono fondamentali per determinare il rischio potenziale di questi organismi ai disturbi antropogenici. Questi risultati saranno utilizzati per informare le future valutazioni del rischio e le strategie di gestione nel quadro della Nuova Zelanda per la gestione dei coralli.
Data Availability Statement
Tutti i set di dati generati per questo studio sono inclusi nell’articolo/materiale supplementare.
Author Contributions
PM, DT, HB, e NH hanno contribuito all’ideazione e alla progettazione dello studio. PM ha eseguito tutta la preparazione dei campioni, l’analisi delle immagini di sezionamento, lo sviluppo del protocollo delle sezioni sottili e il conteggio delle zone, e la micromillatura dei campioni di radiocarbonio. NH ha analizzato i campioni di radiocarbonio. SF ha prodotto stime corrette dell’età dai dati grezzi del radiocarbonio e ha scritto alcune sezioni del manoscritto. PM e DT hanno scritto la prima bozza del manoscritto. Tutti gli autori hanno contribuito alla revisione del manoscritto.
Finanziamento
Questa ricerca è stata finanziata dal secondo anno del Conservation Services Programme (CSP), Progetto POP2017-07 del Dipartimento della Conservazione (DOC) con l’obiettivo di “Sviluppare una metodologia per determinare le caratteristiche di età e crescita delle principali specie di corallo neozelandesi ad alto rischio in acque profonde (fredde)”. Il finanziamento per le spese di pubblicazione di questa rivista è stato fornito da NIWA, Nuova Zelanda.
Conflitto di interessi
Gli autori dichiarano che la ricerca è stata condotta in assenza di relazioni commerciali o finanziarie che potrebbero essere interpretate come un potenziale conflitto di interessi.
Riconoscimenti
Ringraziamo il Conservation Services Programme Group, Department of Conservation New Zealand, in particolare Ian Angus, Kris Ramm, e anche Freydis Hjorvarsdottir (precedentemente del CSP ora Ministry of Primary Industries New Zealand) per il loro supporto al lavoro. Grazie a Helen Neil (NIWA) e Dan Sinclair (VUW), per aver sostenuto la fornitura dei dati preliminari. Ringraziamo il personale NIWA Sadie Mills e Diana Macpherson per il loro contributo nella selezione dei campioni, Owen Anderson per la produzione della Figura 1, Peter Horn per aver fornito un’accurata revisione interna e Rosemary Hurst per utili commenti e consigli sul manoscritto. Ringraziamo Jeremy Horowitz (James Cook University, Australia) per l’identificazione dei campioni, e Dennis Opresko (Emerito Smithsonian Institute) per i commenti sulle revisioni della tassonomia del corallo nero. Infine, ringraziamo i tre revisori, questo documento ha notevolmente beneficiato della loro valutazione dettagliata e costruttiva.
Adkins, J. F., Griffin, S., Kashgarian, M., Cheng, H., Druffel, E. R. M., Boyle, E. A., et al. (2002). Datazione al radiocarbonio di coralli di acque profonde. Radiocarbon 44, 567-580. doi: 10.1017/s0033822200031921
CrossRef Full Text | Google Scholar
Althaus, F., Williams, A., Schlacher, T. A., Kloser, R. J., Green, M. A., Barker, B. A., et al. (2009). Gli impatti della pesca a strascico sugli ecosistemi corallini profondi delle montagne sottomarine sono di lunga durata. Mar. Ecol. Prog. Ser. 397, 279-294. doi: 10.3354/meps08248
CrossRef Full Text | Google Scholar
Alves, E. Q., Macario, K., Ascough, P., and Bronk Ramsey, C. (2018). L’effetto serbatoio radiocarbonico marino mondiale: definizioni, meccanismi e prospettive. Rev. Geophys. 56, 278-305. doi: 10.1002/2017RG000588
CrossRef Full Text | Google Scholar
Anderson, O., Tracey, D., Bostock, H., Williams, M., and Clark, M. (2014). Raffinato Habitat Suitability Modelling per le specie di corallo protette nella ZEE della Nuova Zelanda. NIWA Client Report preparato per il Dipartimento della Conservazione. WLG2014-69No. 46. Auckland: NIWA.
Google Scholar
Andrews, A. H., Cailliet, G. M., Kerr, L. A., Coale, K. H., Lundstrom, C., and DeVogelaere, A. P. (2005). “Investigations of age and growth for three deep-sea corals from the Davidson Seamount off central California,” in Cold-Water Corals and Ecosystems, eds A. Freiwald and J. M. Roberts (Berlin: Springer-Verlag), 1021-1038. doi: 10.1007/3-540-27673-4_51
CrossRef Full Text | Google Scholar
Andrews, A. H., Cordes, E. E., Mahoney, M. M., Munk, K., Coale, K. H., Cailliet, G. M., et al. (2002). Età, crescita e convalida dell’età radiometrica di una gorgonia di acque profonde che forma un habitat (Primnoa resedaeformis) dal Golfo di Alaska. Hydrobiologia 471, 101-110.
Google Scholar
Anthony, K. R. N., and Marshall, P. A. (2009). Le barriere coralline e il cambiamento climatico. Marine Climate Change Impacts and Adaptation Report Card for Australia 2009. Gland: IUCN.
Google Scholar
Bronk Ramsey, C. (2001). Sviluppo del programma di calibrazione radiocarbonica OxCal. Radiocarbon 43, 355-363. doi: 10.1017/s0033822200038212
CrossRef Full Text | Google Scholar
Cairns, S., Gershwin, L., Brook, F., Pugh, P., Dawson, E., Ocanþa, O., et al. (2009). “Phylum cnidaria: coralli, medusae, idroidi myxozo-ans,” in New Zealand Inventory of Biodiversity, Volume 1: Kingdom Animalia, ed. D. P. Gordon (Nuova Zelanda: Canterbury University Press), 59-101.
Google Scholar
Carreiro-Silva, M., Andrews, A. H., Braga-Henriques, A., de Matos, V., Porteiro, F. M., and Santos, R. S. (2013). Variabilità nei tassi di crescita del corallo nero a lungo amato Leiopathes sp. dalle Azzorre. Mar. Ecol. Progr. Ser. 473, 189-199. doi: 10.3354/meps10052
CrossRef Full Text | Google Scholar
Chiswell, S. M., Bostock, H. C., Sutton, P. J. H., e Williams, M. J. M. (2015). Oceanografia fisica dei mari profondi intorno alla Nuova Zelanda: una revisione, New Zealand. J. Mar. Freshw. Res. 49, 1-32. doi: 10.1080/00288330.2014.992918
CrossRef Full Text | Google Scholar
Clark, M., Bowden, D., Baird, S., e Stewart, R. (2010). Effetti della pesca sulla biodiversità bentonica delle montagne sottomarine del complesso “Graveyard”, Chatham Rise settentrionale. Nuova Zelanda. Aqu. Environ. Biodivers. Rep. 46:40.
Google Scholar
Clark, M., and Rowden, A. (2009). Effetti della pesca a strascico in acque profonde sui gruppi di macroinvertebrati delle montagne sottomarine sul Chatham Rise, Nuova Zelanda. Deep Sea Res. I 56, 1540-1554. doi: 10.1016/j.dsr.2009.04.015
CrossRef Full Text | Google Scholar
Clark, M., Tracey, D., Anderson, O., and Parker, S. (2014). Valutazione del rischio ecologico pilota per i coralli protetti. Wellington: National Institute of Water and Atmospheric Research, 32.
Google Scholar
Clark, M. R., Bowden, D. A., Rowden, A. A., e Stewart, R. (2019). Poche prove di resilienza della comunità bentonica alla pesca a strascico su montagne marine dopo 15 anni. Front. Mar. Sci. 6:63. doi: 10.3389/fmars.2019.00063
CrossRef Full Text | Google Scholar
Coleman, D. C. (1991). Tecniche degli isotopi del carbonio. Cambridge, MA: Academic Press.
Google Scholar
Consalvey, M., MacKay, K., and Tracey, D. (2006). Revisione delle informazioni per le specie protette di corallo di acque profonde nella regione della Nuova Zelanda. NIWA Client Report. WLG2006-85. 60. Auckland: NIWA.
Google Scholar
Druffel, E. R. M., Griffin, S., Witter, A., Nelson, E., Southon, J., Kashgarian, M., et al. (1995). Gerardia: pino setoloso del mare profondo? Geochim. Cosmochim. Acta 59, 5031-5036. doi: 10.1016/0016-7037(95)00373-8
CrossRef Full Text | Google Scholar
Druffel, E. R. M., King, L. L., Belastock, R. A., and Buesseller, K. O. (1990). Tasso di crescita di un corallo di mare profondo usando 210Pb e altri isotopi. Geochim. Cosmochim. Acta 54, 1493-1500.
Google Scholar
Fallon, S. J., Fifield, L. K., and Chappell, J. M. (2010). Il prossimo capitolo della datazione al radiocarbonio presso l’Australian National University: relazione sullo stato di avanzamento dell’AMS a stadio singolo. Nuclear Instr. Methods Phys. Res. Sec. B 268, 898-901. doi: 10.1016/j.nimb.2009.10.059
CrossRef Full Text | Google Scholar
Freeman, D., and Cryer, M. (2019). “Current management measures and threats”, in The State of Knowledge of Deep-Sea Corals in the New Zealand Region, Chap. 9, eds D. M. Tracey and F. Hjorvarsdottir (Auckland: NIWA Science and Technology Series).
Google Scholar
Gammon, M. J, Tracey, D. M., Marriott, P. M., Cummings, V. J., e Davy, S. K. (2018). La risposta fisiologica del corallo di profondità Solenosmilia variabilis all’acidificazione dell’oceano. PEERJ 6:e5236. doi: 10.7717/peerj.5236
PubMed Abstract | CrossRef Full Text | Google Scholar
Griffin, S., e Druffel, E. M. (1989). Fonti di carbonio per i coralli di acque profonde. Radiocarbon 31, 533-543. doi: 10.1017/s0033822200012121
CrossRef Full Text | Google Scholar
Hitt, N. T., Sinclair, D. J., Fallon, S. J., Neil, H. L., Tracey, D. M., Komugabe-Dixson, A., et al. (2020). Crescita e longevità dei coralli neri della Nuova Zelanda. Deep Sea Res. I 2020:103298. doi: 10.1016/j.dsr.2020.103298
CrossRef Full Text | Google Scholar
Komugabe, A. F., Fallon, S. J., Thresher, R. E., and Eggins, S. M. (2014). Moderne età del serbatoio di superficie del Mare di Tasmania da coralli neri di profondità. Deep-Sea Res. 99, 207-212. doi: 10.1016/j.dsr2.2013.05.024
CrossRef Full Text | Google Scholar
Komugabe-Dixson, A. F., Fallon, S. J., Eggins, S. M., and Thresher, R. E. (2016). Prove al radiocarbonio per i cambiamenti del medio-tardo Olocene nella circolazione dell’Oceano Pacifico sud-occidentale. Paleoceanography 31, 971-985. doi: 10.1002/2016PA002929
CrossRef Full Text | Google Scholar
Love, M. S., Yoklavich, M. M., Black, B. A., e Andrews, A. H. (2007). Età del corallo nero (Antipathes dendrochristos) colonie, con note sulle specie invertebrate associate. Bull. Mar. Sci. 80, 391-400.
Google Scholar
Matsumoto, K. (2007). Età di circolazione basata sul radiocarbonio degli oceani del mondo. J. Geophys. Res. 112:C09004. doi: 10.1029/2007JC004095
CrossRef Full Text | Google Scholar
Mortensen, P. B., and Buhl-Mortensen, L. (2005). Morfologia e crescita delle gorgonie di acque profonde Primnoa resedaeformis e Paragorgia arborea. Mar. Biol. 147, 775-788. doi: 10.1007/s00227-005-1604-y
CrossRef Full Text | Google Scholar
Noé, S. U., Lembke-Jene, L., e Chr Dullo, W. (2008). Tassi di crescita variabili nei coralli bambù: sclero-cronologia e datazione al radiocarbonio di uno scheletro di gorgonia di acque profonde della metà dell’Olocene (Keratosis sp.: Octocorallia) da Chatham Rise (Nuova Zelanda). Facies 54, 151-166. doi: 10.1007/s10347-007-0129-x
CrossRef Full Text | Google Scholar
Pinkerton, M. H. (2016). Ocean Colour Satellite Observations of Phytoplankton in New Zealand EEZ, 1997-2016. Ministero dell’ambiente. Wellington: NIWA.
Google Scholar
Prouty, N. G., Fisher, C. R., Demopoulous, A. W. J., and Druffel, E. R. M. (2016). Tassi di crescita ed età dei coralli dee-sea impattati dalla fuoriuscita di petrolio della Deepwater Horizon. Deep-Sea Res. II 129, 196-212. doi: 10.1016/j.dsr2.2014.10.021
CrossRef Full Text | Google Scholar
Prouty, N. G., Roark, E. B., Buster, N. A., e Ross, S. W. (2011). Tasso di crescita e la distribuzione dell’età dei coralli neri di acque profonde nel Golfo del Messico. Mar. Ecol. Prog. Ser. 423, 101-115. doi: 10.3354/meps08953
CrossRef Full Text | Google Scholar
Reimer, P. J., Bard, E., Bayliss, A., Beck, J. W., Blackwell, P. G., Bronk Ramsey, C., et al. (2013). IntCal13 e Marine13 curve di calibrazione dell’età radiocarbonica 0-50.000 anni cal BP. Radiocarbon 55, 1869-1887. doi: 10.2458/azu_js_rc.55.16947
PubMed Abstract | CrossRef Full Text | Google Scholar
Reimer, P. J., and Reimer, R. W. (2001). Un database di correzione del serbatoio marino e un’interfaccia online. Radiocarbon 43, 461-463. doi: 10.1017/s0033822200038339
CrossRef Full Text | Google Scholar
Roark, E. B., Guilderson, T. P., Dunbar, R. B., Fallon, S. J., and Mucclarone, D. A. (2009). Estrema longevità in coralli proteinacei di acque profonde. Proc. Natl. Acad. Sci. U.S.A. 106, 5204-5208. doi: 10.1073/pnas.0810875106
PubMed Abstract | CrossRef Full Text | Google Scholar
Roark, E. B., Guilderson, T. P., Dunbar, R. B., and Ingram, B. L. (2006). Età e tassi di crescita basati sul radiocarbonio: Hawaiian coralli d’altura. Mar. Ecol. Progr. Ser. 327, 1-37.
Google Scholar
Sánchez, J., and Rowden, A. (2006). “Octocoral diversity on New Zealand Seamounts,” in Proceedings of the Presented at the 10th International Coral Reef Symposium, Okinawa.
Google Scholar
Sherwood, O. A., and Edinger, E. N. (2009). Età e tassi di crescita di alcune gorgonie di acque profonde e coralli antipatari di Terranova e Labrador. Can. J. Fish. Aquatic Sci. 66, 142-152. doi: 10.1139/f08-195
CrossRef Full Text | Google Scholar
Sherwood, O. A., Scott, D. B., Risk, M. J., and Guilderson, T. P. (2005). Prove al radiocarbonio per anelli di crescita annuali nell’octocoral Primnoa resedaeformis. Mar. Ecol. Prog. Ser. 301, 129-134. doi: 10.3354/meps301129
CrossRef Full Text | Google Scholar
Sikes, E., Burgess, S., Grandpre, R., and Guilderson, T. (2008). Valutare le età moderne delle acque profonde nella regione della Nuova Zelanda utilizzando coralli d’acqua profonda. Deep Sea Res. I 55, 38-49. doi: 10.1016/j.dsr.2007.10.004
CrossRef Full Text | Google Scholar
Stuiver, M., and Polach, H. A. (1977). Discussione sulla segnalazione di dati 14C. Radiocarbon 19, 355-363. doi: 10.1017/S0033822200003672
CrossRef Full Text | Google Scholar
Thresher, R., Rintoul, S. R., Koslow, J. A., Weidman, C., Adkins, J., and Proctor, C. (2004). Prove oceaniche dei cambiamenti climatici nell’Australia meridionale negli ultimi tre secoli. Geophys. Res. Lett. 31:l07212.
Google Scholar
Thresher, R. E., MacRae, C. M., Wilson, N. C., and Gurney, R. (2007). Effetti ambientali sulla composizione scheletrica delle gorgonie d’acqua profonda (Keratoisis spp.; Isididae). Bull. Mar. Sci. 8, 409-422.
Google Scholar
Tracey, D., Bostock, H., and Shaffer, M. (2018). Metodi di invecchiamento per i coralli protetti del mare profondo: A Review and Recommendation for an Ageing Study. Contratto DOC 4527 GMC – Età e crescita del corallo (POP2017-07). NIWA Client Report WN2018035. Auckland: NIWA, 40.
Google Scholar
Tracey, D., Neil, H., Gordon, D., and O’Shea, S. (2003). Cronache del profondo: invecchiamento dei coralli di acque profonde nelle acque della Nuova Zelanda. NIWA Water Atmosph. 11, 22-24.
Google Scholar
Tracey, D. M., e Hjorvarsdottir, F. (eds) (2019). “The state of knowledge of deep-sea corals in the new zealand region,” in NIWA Science and Technology Series Number 84, (Auckland: NIWA),140.
Google Scholar
Tracey, D. M., Neil, H., Marriott, P., Andrews, A. H., Cailliet, G. M., and Sanchez, J. A. (2007). Età e crescita di due generi di coralli di bambù di acque profonde (famiglia Isididae) nelle acque della Nuova Zelanda. Bull. Mar. Sci. 81, 393-408.
Google Scholar
Williams, A., Schlacher, T. A., Rowden, A. A., Althaus, F., Clark, M. R., Bowden, D. A., et al. (2010). Assemblee megabentonici Seamount non riescono a recuperare dagli impatti della pesca a strascico. Mar. Ecol. 31, 183-199. doi: 10.1111/j.1439-0485.2010.00385.x
CrossRef Full Text | Google Scholar
Williams, B., and Grottoli, A. G. (2010). Variabilità degli isotopi stabili dell’azoto e del carbonio (δ15N e δ13C) in coralli molli del Pacifico tropicale poco profondi e taxa di corallo nero e implicazioni per ricostruzioni paleoceanografiche. Geochim. Cosmochim. Acta 74, 5280-5288. doi: 10.1016/j.gca.2010.06.026
CrossRef Full Text | Google Scholar
Williams, B., Risk, M. J., Ross, S. W., and Sulak, K. J. (2007). Dati di isotopi stabili da antipatari di acque profonde: Registrazioni di 400 anni dalla costa sud-orientale degli Stati Uniti d’America. Bull. Mar. Sci. 81, 437-447.
Google Scholar
Leave a Reply