Frontiers in Marine Science
Introduction
Tiefseekorallen sind eine äußerst vielfältige Gruppe von Meeresorganismen, und die Region Neuseeland ist ein Hotspot für Tiefseekorallen (Sánchez und Rowden, 2006; Cairns et al., 2009). In mehreren Studien wurden die Auswirkungen der Grundschleppnetzfischerei auf die Makroinvertebratengemeinschaften der Tiefsee (einschließlich Korallen) im Südpazifik untersucht (Althaus et al., 2009; Clark und Rowden, 2009; Clark et al., 2010, 2019; Williams et al., 2010; Anderson et al., 2014). Diese haben gezeigt, dass die Grundschleppnetzfischerei spürbare Auswirkungen auf langsam wachsende und empfindliche Tiefseekorallenkolonien und -gemeinschaften hat, die sich kaum erholen können. Die durch den Klimawandel vorhergesagten Umwelteinflüsse wie die Erwärmung der Ozeane und die Versauerung der Meere werden voraussichtlich auch die Gesundheit der Tiefseekorallen bedrohen, da einige Lebensräume zu warm werden, damit sie gedeihen können (Anthony und Marshall, 2009), oder der pH-Wert und die Karbonat-Ionen sinken, was sich wahrscheinlich auf Tiefseekorallen mit Karbonat-Skeletten auswirken wird (Gammon et al., 2018). Um künftige Risikobewertungen und die Bewirtschaftung der Meeresressourcen besser zu informieren, ist das Verständnis des Alters und des Wachstums von Tiefseekorallen der Schlüssel zur Bestimmung der Regenerationszeiten und der Erholungsfähigkeit von Korallen nach anthropogenen Störungen oder anderen natürlichen Schäden.
In der Region Neuseeland wurde eine beträchtliche Menge an Arbeiten zur Taxonomie von Tiefseekorallen, zur Verbreitung verschiedener Arten und ihrer Umweltpräferenzen sowie zur Bedeutung der Tiefseekorallen für die benthischen Ökosysteme durchgeführt. Vor kurzem wurde ein Überblick über unser Verständnis und Wissen über Tiefseekorallen in der Region Neuseeland veröffentlicht (Tracey und Hjorvarsdottir, 2019). In dieser Übersicht wurden auch große Lücken in unserem Verständnis von Tiefseekorallen aufgezeigt, einschließlich begrenzter Alters- und Wachstumsdaten für viele Gattungen und Arten. In dieser Studie bewerten wir das Alter und die Wachstumsrate der geschützten schwarzen Tiefseekoralle Antipatharian Bathypathes patula. Das Alter und die Wachstumsrate dieser Koralle wurden in dieser Region bisher noch nicht untersucht. Die Auswahl dieser Art basierte auf der Prioritätenliste der Risikobewertung (Clark et al., 2014) und der Literaturübersicht (Tracey et al., 2018), gepaart mit der Verfügbarkeit von Proben (Standort und Gesamtzahl) und anderen ergänzenden Forschungen zu schwarzen Korallen, die in Neuseeland durchgeführt werden (Hitt et al., 2020).
Tracey et al. (2018) skizzierten die aktuellen Methoden, die zur Altersbestimmung von Tiefseekorallenarten verwendet werden. Die Vor- und Nachteile der einzelnen Methoden wurden diskutiert. Die wichtigsten Methoden, die erfolgreich zur Messung des Alters und des Wachstums von Tiefseekorallen angewandt wurden, sind (1) direkte Beobachtung, wie z. B. In-situ-Messungen oder In-aquaria-Experimente zum linearen Wachstum oder zu Oberflächenausdehnungen; Polypenadditionsrate; Schätzung der Kalzifizierungsraten (z. B., (2) Auszählung von Skelettwachstumszonen oder -bändern (Sklerochronologie) und (3) radiometrische Analysen.
Direkte Beobachtungen und In-situ- oder In-aquaria-Messungen sind bei Tiefseekorallen sehr schwierig durchzuführen, da es schwierig ist, lebende, lebensfähige Exemplare zu erhalten und sie am Leben zu halten, da sie eine lange Zeit für ihr Wachstum benötigen (Gammon et al., 2018; Tracey et al., 2018). Die Aufzählung von Wachstumszonen, die Sclerochronologie, ist ideal für Tiefseekorallenarten, die einen hohen Kontrast zwischen den zonenartigen Wachstumsmustern aufweisen, die in Skelettquerschnitten beobachtet werden. Es ist jedoch unerlässlich, dass eine unabhängige Validierungsmethode verwendet wird, um die chronologische Gültigkeit der Zählungen der beobachteten Wachstumszonen zu prüfen (Andrews et al., 2002). Die Methode zur Zählung der sichtbaren Wachstumszonen in der Skelettstruktur wurde erfolgreich angewandt, um das Alter und die Wachstumsrate von Oktokorallen zu bestimmen, darunter die 400 Jahre alte Bambuskoralle Keratoisis sp, 100 Jahre alte Seefische Primnoa spp. und die 60 Jahre alte Bambuskoralle Lepidisis sp. (Thresher et al., 2004, 2007; Mortensen und Buhl-Mortensen, 2005; Sherwood et al., 2005; Tracey et al., 2007; Sherwood und Edinger, 2009). Schwarze Korallen wurden auch erfolgreich anhand von Wachstumszonen oder Zählungen gealtert, mit einem Alter von 150 bis 480 Jahren für Antipathes dendrochristos bzw. Leiopathes glaberrima (Love et al., 2007; Williams et al., 2007). In einer Studie über die Schwarze Koralle Stauropathes arctica wurden Zonenzahlen von 55-58 ermittelt (Sherwood und Edinger, 2009), wobei die Autoren anmerkten, dass die Radiokohlenstoffdatierung diese Alter auf 55 und 82 Jahre einschränkte. Die Wachstumsraten schwarzer Korallen durch Aufzählung der Wachstumszonen zeigen ein geringes radiales Wachstum von 8 bis 140 μm/Jahr (Love et al., 2007; Prouty et al., 2011).
Es gibt potenzielle Einschränkungen bei der Aufzählung der Wachstumszonen (siehe Zusammenfassung in Tracey et al., 2018). Viele Korallen weisen keine klare regelmäßige Zonenstruktur in ihrem gesamten Skelett auf, oder die klare Zonenstruktur ist nur in Teilen des Skeletts erkennbar. Selbst wenn sie vorhanden ist, kann die Zonenstruktur variabel sein oder unterschiedliche Interpretationsmöglichkeiten bieten. So waren beispielsweise die Basal- und Spitzenzahlen der Bambus-Krakenarten Keratoisis sp. und Lepidisis sp. höher als die mit der radiometrischen Methode Blei 210 (210Pb) geschätzten Alter (Tracey et al., 2007). Es wurde die Hypothese aufgestellt, dass die mit dem Lichtmikroskop beobachteten Zonen eine halbjährliche Periodizität aufweisen und dass die mit dem REM beobachteten Zonen an den Knotenpunkten ein Umweltereignis, z. B. eine Mondperiode, darstellen könnten. Mondzyklen können mit starken Gezeitenereignissen verbunden sein, die das Nahrungsangebot für sessile, filtrierende Organismen verbessern und zu somatischen Wachstumsschüben führen könnten. Gorgonien (z. B. Paragorgia spp.) und rote Edelkorallen (z. B. Corallium sp.) wurden aufgrund ihrer komplexen oder schlecht definierten Bändermuster nicht erfolgreich mit Hilfe von Zählungen gealtert (Griffin und Druffel, 1989; Andrews et al, 2005).
Die am häufigsten verwendete radiometrische Methode bei Tiefseekorallen ist die Datierung mit dem Isotop Radiokohlenstoff (14C) (Druffel et al., 1995; Sherwood et al., 2005; Tracey et al., 2018). Diese Methode beruht auf der Tatsache, dass eine sehr geringe Menge des natürlichen Kohlenstoffs in der Atmosphäre in Form von 14C radioaktiv ist und in das Korallenskelett eingebaut wird, wenn es sein Kalziumkarbonat oder Protein und Chitin bildet (Adkins et al., 2002; Tracey et al., 2003; Consalvey et al., 2006). Da die Halbwertszeit von Radiokohlenstoff bekanntlich 5.730 Jahre beträgt, kann diese Methode, auch wenn sie teuer ist, verwendet werden, um Proben, die weniger als 50.000 Jahre alt sind, zuverlässig zu altern; darüber hinaus wird die Aktivität von 14C zu gering, um sie nachzuweisen (Coleman, 1991). In den 1950er- und 1960er-Jahren kam es zu einem raschen Anstieg des atmosphärischen 14C infolge der Tests von Nuklearsprengkörpern; dieser „Bombenkohlenstoff“ kann ebenfalls zur Kalibrierung von Altersangaben verwendet werden und bietet einen Bezugspunkt für jüngere Proben (Coleman, 1991; Druffel et al., 1995; Tracey et al., 2003; Roark et al, 2009; Sherwood und Edinger, 2009).
Die Radiokohlenstoffdatierung wurde bei mehreren Korallenarten, darunter Steinkorallen, Schwarzkorallen und Gorgonien, erfolgreich eingesetzt (Druffel et al., 1990; Roark et al., 2006; Carreiro-Silva et al., 2013; Prouty et al., 2016; Hitt et al., 2020). Allerdings gibt es auch bei dieser Methode Einschränkungen. Infolge der Ozeanzirkulation ist das Radiokohlenstoffalter des Wassers in den Ozeanen deutlich älter als das der Atmosphäre, das so genannte „marine Reservoiralter“ (Tracey et al., 2003; Consalvey et al., 2006; Roark et al., 2006; Noé et al., 2008). Fehler im Alter des Reservoirs können erhebliche Auswirkungen auf die Schätzung des Korallenalters haben. Die Korrektur des Reservoir-Alters erfordert Kenntnisse über die umgebenden Gewässer und geht von konstanten Radiokohlenstoffverhältnissen im Wasser aus; diese Werte können jedoch durch Vermischung im Ozean, lokale Auftriebsereignisse und Produktivitätsblüten verändert werden, die alle die Kohlenstoffkreislaufumwandlungen und die 14C-Zusammensetzung des Korallenskeletts verändern können (Tracey et al, 2018).
Basierend auf der Übersichtsarbeit von Tracey et al. (2018) haben wir uns für die Anwendung von zwei Alterungsmethoden für B. patula entschieden: die Zählung von Skelett-Wachstumsstreifen und die radiometrische Methode der Radiokohlenstoffdatierung (14C). Die Anwendung dieser Methoden ermöglicht Vergleiche mit anderen früheren Korallenalterungsstudien sowohl in Neuseeland als auch weltweit. Die Probenauswahl, das Mikrofräsen des Materials und die Schnittmethoden sowie die Interpretation der Zonenzählung und der Radiokohlenstoffdatierung werden vorgestellt. Alter und Wachstumscharakteristika der schwarzen Koralle B. patula und einer Kolonie von Bathypathes cf. conferta (ursprünglich als B. patula angenommen) werden dargestellt.
Materialien und Methoden
Probenauswahl
Die in dieser Studie verwendeten Proben stammen von konservierten Exemplaren, die im neuseeländischen National Institute of Water and Atmospheric Research (NIWA), National Invertebrate Collection (NIC), gelagert sind.
Proben wurden aus zwei Regionen ausgewählt: der Bay of Plenty (nordöstlich der Nordinsel Neuseelands) und dem Chatham Rise (östlich von Neuseeland), um die Ergebnisse zu vergleichen und die Variabilität der Wachstumsraten zwischen den verschiedenen Gebieten zu bewerten, und auch mit anderen Tiefsee-Schwarzkorallengattungen, insbesondere Antipathella, Leiopathes und Antipathes, aus denselben Regionen (Hitt et al., 2020).
Ozeanografische Strömungen beeinflussen Tiefseekorallen, indem sie für einen stetigen Strom von Nahrung und die Verbreitung von Larven sorgen. Folglich beeinflussen diese Strömungen auch die Verfügbarkeit von Beutetieren und die Verbreitung von Tiefseekorallen. Während Tiefseekorallen am meisten von den Bodenwasserströmungen beeinflusst werden, die über sie hinwegfließen und die Verteilung der Sedimente beeinflussen, werden sie auch von den Oberflächenwasserströmungen beeinflusst, die die Primärproduktivität und damit das Nahrungsangebot beeinflussen, das auf die Tiefseekorallen am Meeresboden niederregnet. Die neuseeländische Landmasse liegt an der Schnittstelle zwischen warmen, nährstoffarmen, subtropischen Oberflächengewässern, die von Norden her einströmen, und kühlen, nährstoffreichen, subantarktischen Oberflächengewässern, die von Süden her einströmen. Das derzeitige Verständnis der Wassermassen und Meeresströmungen in der Region an der Oberfläche, in der Sprungschicht, im Zwischenwasser und im Tiefenwasser wurde kürzlich von Chiswell et al. (2015) überprüft. Die ozeanografischen Bedingungen für die Korallen aus dieser Studie sind wie folgt:
Die Bay of Plenty wird von warmen, nährstoffarmen, subtropischen Oberflächengewässern umspült, die aus dem ostaustralischen Strom stammen und von Westen her einströmen und den East Auckland Current bilden. Am Chatham Rise treffen die warmen, nährstoffarmen, subtropischen Gewässer (STW) und die kühlen, nährstoffreichen, subantarktischen Gewässer (SAW) aus dem Südlichen Ozean an der Subtropischen Front (STF) aufeinander. Die Vermischung von STW- und SAW-Wassermassen an der STF führt dazu, dass der Chatham Rise eine Zone mit sehr hoher Primärproduktivität ist, die eine bedeutende Nahrungsquelle und damit Kohlenstoffversorgung für diese Korallen darstellt. Die Chlorophyll-a-Konzentrationen liegen zwischen 0,3 und 0,48 mg/m3 im Chatham Rise und in der Bay of Plenty, verglichen mit 0,03-0,3 mg/m3 im zentralen Pazifik (Pinkerton, 2016). In Tiefen von 500-1500 m strömen die kühlen (3,5-10°C), salzarmen (34,4-34,6) und sauerstoffreichen (200-250 μmol/kg) sowie nährstoffreichen (Nitrat 15-36 μmol/kg) antarktischen Zwischenwässer (AAIW) aus dem Norden und Süden in diese Region (Chiswell et al, 2015).
Korallen aus dem Chatham Rise wurden ausgewählt, da es moderne 14C-Reservoir-Altersdaten aus diesem Gebiet gibt (Sikes et al., 2008). Die Kenntnis des lokalen 14C-Reservoiralalters ermöglicht eine präzisere Radiokohlenstoff-Altersschätzung von Korallen.
Die Proben wurden aus vorhandenen Exemplaren ausgewählt, die von Beobachtern der Fisheries New Zealand (FNZ), des Ministry for Primary Industries (MPI) und von NIWA-Forschern gesammelt wurden und im (NIC) aufbewahrt werden. Eine Darstellung der Verbreitungsdaten von B. patula half bei der Entscheidung über die Auswahl der Kolonie und die Anzahl der Proben (Abbildung 1). Zwei Exemplare aus der Bay of Plenty (NIWA24190 und NIWA85940) wurden für die Radiokarbondatierung und Dünnschliffe für die Zählung der Zonen ausgewählt. Die übrigen Exemplare wurden von der Chatham Rise ausgewählt.
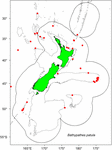
Abbildung 1. Standorte, an denen Bathypathes patula in der Region Neuseeland beprobt wurde. Die roten Punkte bezeichnen die Fundorte einzelner Korallenproben von B. patula, die in der National Invertebrate Collection of New Zealand (NIC) aufbewahrt werden. Graue Linien kennzeichnen die 200 m Wassertiefenkontur. Die schwarze Linie kennzeichnet die Ausdehnung der Ausschließlichen Wirtschaftszone (AWZ) Neuseelands.
Ende 2018 überprüfte ein Gastspezialist für die Taxonomie schwarzer Korallen (Jeremy Horowitz, James Cook University) die Identifizierungen mehrerer Proben in der NIC. Während dieses Prozesses wurde eine der schwarzen Korallenkolonien, die für diese Altersbestimmung ausgewählt wurde und von der angenommen wurde, dass es sich um Bathypathes patula handelt, einer neuen Identifizierung unterzogen. Die schwarze Koralle NIWA24190 wurde nun als Bathypathes cf. conferta identifiziert. Die Probe wird in diesem Dokument durchgehend als Bathypathes cf. conferta bezeichnet (Abbildung 2), was die wahrscheinlichste korrekte Identifizierung darstellt. Eine weitere taxonomische Überprüfung dieses Exemplars ist im Gange (Opresko pers comm). Die Verbreitungskarte (Abbildung 1) kann weitere Exemplare von B. cf. conferta enthalten.

Abbildung 2. Bathypathes cf. conferta. NIWA24190. Das Exemplar zeigt, wo die Proben für die Analyse entnommen wurden. (A) ist die wachsende Spitze, (B) der Basalschnitt. Exemplar aus der Bay of Plenty, Neuseeland. Maßstabsleiste ist ein 35-cm-Lineal.
Materialvorbereitung
Nachdem geeignete Proben identifiziert worden waren, wurden kleine (weniger als 10 mm große) Abschnitte aus den Wachstumsspitzen und dem basalen Teil des Hauptstamms der Korallen entnommen. Da viele der Proben während der Entnahme gebrochen waren, wurden zusätzliche Abschnitte entlang des Hauptstammes entnommen, damit zuverlässige Schätzungen der linearen Wachstumsrate erstellt werden konnten (Abbildungen 2-5). Die Proben von den Wachstumsspitzen wurden dann in zwei Fragmente aufgeteilt, eines für die Radiokohlenstoffdatierung, das andere für die Präparation von Dünnschliffen, um Zonenzahlen zu erhalten.

Abbildung 3. Bathypathes patula NIWA85940. Das Exemplar zeigt, wo die Proben für die Analyse entnommen wurden. (A) ist die Wachstumsspitze, (C) der basale Abschnitt und (B) ist eine Zwischenprobenstelle, da das Exemplar bei der Entnahme gebrochen war und unbekannte Längen des Hauptstammes oberhalb dieser Stelle fehlen könnten. Exemplar aus der Bay of Plenty, Neuseeland. Maßstabsleiste ist ein 35 cm Lineal.

Abbildung 4. Bathypathes patula NIWA49468. Das Exemplar zeigt, wo die Proben für die Analyse entnommen wurden. (A) ist die Wachstumsspitze, (D) der Basalschnitt, und (B,C) sind Zwischenproben, da das Exemplar bei der Entnahme gebrochen war und eine unbekannte Länge des Hauptstammes zwischen diesen beiden Stellen fehlen könnte. Exemplar von Chatham Rise, Neuseeland. Maßstabsleiste ist ein 35 cm Lineal.

Abbildung 5. Bathypathes patula NIWA47911. Das Exemplar zeigt, wo die Proben für die Analyse entnommen wurden. (A) ist die wachsende Spitze, (F) der basale Abschnitt und (B-E) sind Zwischenproben, die entnommen wurden, um das Radiokarbonsignal zu suchen. Exemplar von Chatham Rise, Neuseeland. Der Maßstab ist ein 35 cm langes Lineal.
Mikrofräsen des Materials
Bei früheren Arbeiten wurde festgestellt, dass schwarze Korallenabschnitte dazu neigen, durch Kapillarwirkung Harz durch ihre poröse Matrix nach oben zu ziehen, so dass die Abschnitte in dieser Studie nicht mit Harz auf eine Grundplatte zum Fräsen geklebt werden konnten. Stattdessen wurde ein Spannfutter hergestellt, um den Abschnitt während des Fräsens zu halten und so das Risiko einer Verunreinigung der Radiokohlenstoffproben durch Harz auszuschalten.
Pulverisierte Radiokohlenstoffproben wurden aus dem Rand und dem Kern von Basal-, Mittel- und Spitzenabschnitten mit einer New WaveTM-Mikromühle mit einem 0,5 mm Brassler H2.11.006-Fräsbohrer am NIWA entnommen. Die Randproben umfassten Material aus den äußeren 250 Mikrometern des Abschnitts, die Kernproben wurden aus Material innerhalb eines Radius von 750 Mikrometern um das Primordium gewonnen. Wir versuchten, mindestens 1,5 mg Material pro Probe zu gewinnen. Die Proben wurden dann gewogen und im Radiokohlenstofflabor der Australian National University (ANU) auf Radiokohlenstoff analysiert.
Dünnschnittmethode und Zählprotokolle
Korallenstammabschnitte wurden in durchsichtiges, zweiteiliges Epoxidharz eingebettet und am NIWA mit einer Diamantsäge geschnitten. Die Schnitte wurden auf einer Seite poliert und mit der polierten Seite nach unten auf einen Glasmikroskop-Objektträger montiert, dann geschliffen und poliert, bis sie eine optimale Dicke für die Betrachtung der Wachstumszonenstruktur im Durchlicht hatten. Die optimale Dicke für Schnitte dieser Art betrug etwa 250-300 μm und entspricht dem Standardverfahren für die Dünnschnittpräparation (Andrews et al., 2002; Mortensen und Buhl-Mortensen, 2005; Tracey et al., 2007). Die Zählung der periodischen Wachstumsbänder oder Inkremente, von denen angenommen wird, dass sie jährlich gebildet werden, erfolgte nach der Dünnschliffpräparation am NIWA.
Um die Zählung der Zonen zu erstellen, wurden Dünnschliffpräparate der Hauptastabschnitte unter einem Verbundmikroskop mit Durchlicht betrachtet. Als Hilfsmittel für die Zoneninterpretation wurden die Schnitte auch mit ultraviolettem Licht nach der Methodik von Sherwood und Edinger (2009) betrachtet. In diesem Fall wurden die Zonen mit durchscheinendem Hellfeldlicht gezählt, und die ultraviolette Beleuchtung war ein nützliches Hilfsmittel, um die beobachtete Zonenstruktur weiter zu definieren.
Radiokohlenstoff-Datierungsmethode
Vor der Analyse wurden die Proben von schwarzen Krusten und endolithischen Aktivitäten gereinigt (z. B. durch Säureauslaugung), um potenzielle jüngere Verunreinigungen mit 14C zu entfernen, die die Ergebnisse verfälschen könnten (Adkins et al., 2002). Die pulverisierten Korallenproben wurden mit <1 ml 0,1 M HCL behandelt. Die Säure wurde dann abpipettiert und die Proben wurden dreimal mit milliQ-Wasser gewaschen. Anschließend wurde die Probe mit Trockeneis eingefroren und gefriergetrocknet. Etwa 1 mg der gereinigten Korallenprobe wurde in ein Quarzrohr mit 6 mm OD eingewogen, ∼60 mg CuO und ein Silberbecher wurden hinzugefügt. Die Röhren wurden auf <3e-3 Torr (<0,4 Pa) evakuiert und mit einer Taschenlampe versiegelt. Die Röhren wurden dann 6 Stunden lang bei 900 °C gebacken, um CO2 zu erzeugen. Das entstandene CO2 wurde gereinigt und in Anwesenheit von Wasserstoff mit angetriebenem Fe als Katalysator in Graphit umgewandelt. Der Graphit wurde dann mit dem einstufigen Beschleuniger-Massenspektrometer der ANU gemessen (Fallon et al., 2010). Alle Daten wurden mit Online-AMS δ13C korrigiert, auf Oxalsäure I normalisiert und der Hintergrund mit 14C-freier Kohle subtrahiert, die auf die gleiche Weise wie die Korallenproben behandelt wurde. Die Daten werden gemäß den Empfehlungen von Stuiver und Polach (1977) dargestellt.
Ergebnisse
Dünnschliffe für Zonenzählungen
Die an den Skelettschnitten dieser Arten beobachteten Zonenstrukturmuster sind sehr komplex und können auf verschiedene Weise interpretiert werden. Für die Zählung des Radialwachstums wurden die präparierten Schnitte sorgfältig beobachtet, und die Achse, die am besten eine klare Zonenstruktur vom Primordium bis zum äußeren Rand aufwies, wurde dann für die Zählung der Zonen verwendet. Das radiale Wachstum der Zweige von B. patula verlief einigermaßen regelmäßig, und es gab keine ausgeprägte Tendenz zur Bildung dickerer Zonen auf einer bestimmten Seite des Zweigs, wie sie zuvor bei anderen schwarzen Korallenarten beobachtet wurde. Die für die Berechnungen der Wachstumsrate verwendeten Radien waren der Mittelwert der minimalen und maximalen Radien, die in den präparierten Abschnitten beobachtet wurden, was bedeutet, dass die angegebenen Radialwachstumsraten eine mittlere Radialwachstumsrate für die Koralle darstellen.
Auf dem Korallenskelett gibt es ein anfängliches grobes Bindenmuster, das weitgehend durch abwechselnd dunklere und hellere Zonen definiert ist (Abbildung 6), wenn man es mit geringerer Leistung (10-20×) betrachtet. Bei näherer Betrachtung unter höherer Vergrößerung (100-200×) ist auch ein recht regelmäßiges, feines Bänderungsmuster erkennbar (Abbildung 7). Hier haben wir zwei Leseprotokolle angewandt – wir zählten sowohl die grob geformten breiten Zonen als auch die dünnen, fein geformten Zonen (Tabelle 1). Die hier vorgestellten Zählungen wurden alle von einem einzigen Ableser (Marriott) durchgeführt. Während der Entwicklung der Protokolle für die Zonenauswertung wurden viele Ablesungen unter Anwendung der verschiedenen Iterationen der entwickelten Protokolle durchgeführt. Die Zonenzahlen der beiden hier vorgestellten Protokolle wurden jeweils aus mehreren Zählungen im gesamten Abschnitt abgeleitet.

Abbildung 6. Basalschnitt der Koralle NIWA47911, der die grobe farbige Bänderung der beobachteten Zonenstruktur zeigt. Das Exemplar zeigt 54 Zonen, die mit roten Punkten markiert sind. Das gestrichelte Rechteck zeigt den zentralen Teil des in Abbildung 7 abgebildeten Basalschnitts.

Abbildung 7. Zentraler Teil des Basalschnitts der Koralle NIWA47911, der das feinmaßstäbliche Interpretationsprotokoll der beobachteten Zonenstruktur zeigt. Das Exemplar zeigt 42 mit schwarzen Punkten markierte Zonen in diesem inneren Bereich des Schnittes; der gesamte Schnitt zeigte 211 Zonen.

Tabelle 1. Zonenzahlen aus Dünnschliffen.
Für den jüngsten Teil der untersuchten Proben, die Spitze oder die Region nahe der Spitze, lagen die Zählungen zwischen 4 und 7 (grobe Zonenzahlen) und zwischen 10 und 19 (feine Zonenzahlen). In der mittleren Region des Hauptastes lagen die Zählungen zwischen 55 und 60 (Grobzonenzählungen) und 370 und 374 (Feinzonenzählungen). In der Basisregion der Kolonie reichten die Zählungen von 37 bis 193 (Grobzonenzählungen) und von 78 bis 481 (Feinzonenzählungen). Die Komplexität der Zonierungsmuster machte die Notwendigkeit einer Validierung deutlich.
Radiokohlenstoffanalyse
Die in Tabelle 2 aufgeführten gefrästen Proben wurden zur Radiokohlenstoffdatierung an die ANU geschickt. Die Ergebnisse der 17 Korallenproben sind in den Radiokohlenstoffergebnissen (Tabelle 3) aufgeführt. Drei Proben (Proben 47911-F2; 24190-B1; 85940-B1) fielen im Labor aus, da nicht genügend Material für einen Duplikatversuch vorhanden war. Die Radiokohlenstoffergebnisse der verbleibenden Korallen wurden mit OxCal4.3 (Bronk Ramsey, 2001), der Marine13-Kurve und einem lokalen neuseeländischen deltaR (Radiokohlenstoff-Reservoir-Offset) von -18 ± 36 (Reimer und Reimer, 2001) auf ein Kalenderalter kalibriert. Die Altersverteilungen der Korallenproben aus der Zeit vor der Bombardierung sind in Tabelle 3 dargestellt. Alle Korallenproben, die in dieser Studie verwendet wurden, wurden lebend gefangen, so dass alle angegebenen Altersschätzungen das Alter bei der Gefangennahme und nicht die Lebensspanne des Exemplars wiedergeben. Die voraussichtliche Lebensdauer dieser Art wird die hier angegebenen Altersschätzungen übersteigen. Die Koralle 85940 war etwa 380 ± 50 Jahre alt, die Koralle 49468 etwa 310 ± 70 Jahre und die Koralle 47911 etwa 120 ± 50 Jahre (Tabelle 3). Alle Proben der Korallenaußenränder und -spitzen enthielten Radiokarbonwerte F14C >1 (Tabelle 3), mit Ausnahme von 85940-B2 und 85940-C2, was darauf hinweist, dass diese Korallen zum Zeitpunkt der Entnahme lebendig waren und Kohlenstoff aus dem Oberflächenwasser aufnahmen. Die Korallen 85940-B2 und 85940-C2 wiesen am äußeren Rand Radiokohlenstoffwerte F14C <1 (Tabelle 3) auf, was darauf hindeutet, dass ein Teil des Skeletts nicht aktiv Material aufnahm. Dies wurde schon früher beobachtet, wenn der basale Teil der Korallenkolonie aufhört zu wachsen, der obere Teil jedoch weiter wächst (Komugabe-Dixson et al., 2016).
Tabelle 2. Für die Radiokohlenstoffdatierung entnommene Proben.

Table 3.
Schätzungen der Wachstumsraten
Die Schätzungen der Wachstumsraten basierten auf den Ergebnissen der Radiokohlenstoffdatierung, da die Zählung der Zonen problematisch war und sich als nicht validiert erwies, so dass keine zuverlässigen Schätzungen der Wachstumsraten möglich waren (siehe Abschnitt „Bathypathes patula Alters- und Wachstumsschätzungen“ für eine weitere Diskussion über die Zählung der Zonen). Die Wachstumsraten wurden aus den physischen Entfernungen zwischen den Probenahmestellen und den kalibrierten Radiokohlenstoff-Altern an der jeweiligen Probenstelle abgeleitet. Da einige Proben während der Radiokohlenstoffanalyse ausfielen, konnte nur ein begrenzter Satz von Wachstumsratenschätzungen erstellt werden. Die radialen Wachstumsraten, d. h. die Rate, mit der sich der Koloniezweig im Laufe der Zeit radial verdickt, wurden geschätzt, indem die kalibrierten Kalenderjahre des Wachstums zwischen dem Kern und dem Rand der Probenstelle durch den mittleren Radius des Zweigsegments geteilt wurden. Das radiale Wachstum reichte von 11,1 bis 35,7 μm/Jahr. Die linearen Wachstumsraten, d. h. die Rate, mit der sich der Koloniezweig im Laufe der Zeit in Längsrichtung verlängert, wurden geschätzt, indem die Differenz zwischen den kalibrierten Kalenderjahren der Kernproben durch die lineare Entfernung zwischen zwei Probenstellen geteilt wurde. Es wurden zwei lineare Wachstumsraten ermittelt, 5,2 und 9,6 mm/Jahr (Tabelle 4).

Table 4. Bathypathes patula Wachstumsraten-Schätzungen.
Diskussion
Bathypathes patula Alters- und Wachstumsschätzungen
Die Radiokarbon-Ergebnisse dieser Arbeit zeigen, dass B. patula eine langlebige Art ist, mit einem Alter von über 385 Jahren. Langsame lineare (5,2-9,6 mm/Jahr) und radiale (11,1-35,7 μm/Jahr) Wachstumsraten wurden aus den Radiokohlenstoff-Altersdaten gewonnen.
Keines der Zählprotokolle korrelierte zuverlässig mit den unabhängigen Radiokohlenstoff-Altersschätzungen. Das feine Zählprotokoll für die Zonen zeigte die engste Korrelation mit den 14C-Altersdaten, was insgesamt eine gewisse Unterstützung für die Langlebigkeit bietet, wenn man die Zählungen mit den 14C-Daten vergleicht. Die Altersschätzungen zwischen den Radiokohlenstoff-Altersdaten und den feinskaligen Zonenzählungsdaten zeigten routinemäßig eine Diskrepanz von mehr als hundert Jahren in Zweigabschnitten mit kalibrierten Radiokohlenstoff-Altern von 112-385 Jahren (Tabellen 1, 3). Die Unterschiede in den geschätzten Altersangaben von vergleichbaren Proben unter Verwendung der beiden Methoden waren negativ und positiv; es gab keine Unter- oder Überalterung unter Verwendung von Zonenzählungen um einen proportionalen Betrag (Tabelle 1). Das bedeutet, dass die beobachtete Zonenstruktur weder ein jährliches noch ein anderes regelmäßiges chronologisches Muster der Periodizität aufweist, und unsere Zählprotokolle können nicht verwendet werden, um verlässliche Altersschätzungen für B. patula zu erstellen.
Die Rand-Radiokohlenstoff-Ergebnisse des Exemplars NIWA-85940 deuten darauf hin, dass der basale Abschnitt (C2) des Hauptstammes 293 Jahre vor der Ernte aufhörte, Skelettmaterial auf seiner äußeren Oberfläche abzulagern, und der mittlere Abschnitt (B2) hörte 88 Jahre vor der Ernte auf, Skelettmaterial abzulagern. Und das, obwohl diese Abschnitte des Aststammes unmittelbar an die seitlichen Fiederblättchen mit lebenden Polypen angrenzten. Das optische Erscheinungsbild dieser Abschnitte gab keinen Hinweis darauf, dass die Ablagerung von Zonen an ihrem äußeren Rand aufgehört hatte. Die Oberfläche des Korallenastes schien noch relativ unberührt zu sein, sie wies keine Anzeichen von oberflächlicher Erosion oder Verfärbung auf, wie es bei einem Korallenast zu erwarten wäre, der an seiner Außenfläche kein neues Skelettmaterial mehr anlagert. Dies deutet darauf hin, dass, wenn die Stämme der Zweige aufhören können, Skelettschichten zu bilden, während die Zweigenden weiter wachsen, die Protokolle der Zählmethode nicht zuverlässig für die Erstellung von Altersschätzungen aus Basalschnitten für diese Korallenart verwendet werden können.
In der Probe NIWA47911 wurden fünf Proben von der wachsenden Spitze des axialen Hauptastes in Richtung der Korallenbasis entnommen. Diese Proben wurden in einem Abstand von 25 mm entnommen. Auf diese Weise wurde versucht, das Auftreten von Bomben-Radiokohlenstoff im Korallenskelett zu lokalisieren. Dies würde bedeuten, dass die lineare Wachstumsrate für die jüngeren Wachstumsspitzen der Äste der Kolonie genau bewertet werden könnte. Wenn der Anstieg des Bomben-Radiokohlenstoffs genau lokalisiert werden könnte, dann würde dies auch einen genauen Anknüpfungspunkt für die Festlegung der 14C-Korrektur liefern, die auf die Radiokohlenstoffproben aus der Zeit vor der Bombardierung für diese Kolonie und für andere Kolonien aus derselben Wassermasse anzuwenden ist. Leider erwiesen sich alle diese Proben als Nachbombenproben, so dass dieser Prozess nicht abgeschlossen werden konnte. Zukünftige Probenahmen weiter hinten in diesem Zweigstamm wären nützlich, um diesen Verbindungspunkt und die zeitgenössische lineare Wachstumsrate zu bestimmen. Bei unserer Arbeit wurden lineare Gesamtwachstumsraten von 5,2-9,6 mm/Jahr gemessen. Wenn man von der schnellsten Wachstumsrate ausgeht, müsste man von der Wachstumsspitze aus 700 mm zurück in den Hauptast beproben, um sicher zu sein, dass man den Anstieg des Bomben-Radiokohlenstoffs im Skelett beobachten kann.
Forschungen von Love et al. (2007) an der schwarzen Korallenart Antipathes dendrochristos berichteten ebenfalls über zwei Interpretationen der Zonenstruktur, die den hier beschriebenen Grob- und Feinprotokollen ähneln. Diese Autoren fanden jedoch heraus, dass die gröbere farbige Bandstruktur mit ihren Radiokohlenstoff- und 210Pb-Ergebnissen korrelierte, was auf eine jährliche Bandstruktur hindeutet (Love et al., 2007).
Die Radiokohlenstoffmessungen aus der aktuellen Arbeit zeigten F14C >1 in Proben von den wachsenden Astspitzen und der Oberfläche des Hauptastes, was darauf hindeutet, dass diese Proben moderne Radiokohlenstoffwerte aus der Zeit nach der Bombardierung aufweisen. Das Vorhandensein von Nachbombenkohlenstoff unterhalb der gemischten Schicht des Ozeans deutet darauf hin, dass die schwarzen Korallen von B. patula kürzlich exportierten Oberflächenwasser-POC nutzen. In diesen Tiefen ist POC die einzige Kohlenstoffquelle, die mit Bomben-14C markiert ist. Der gelöste anorganische Kohlenstoff (DIC) des Umgebungswassers, in dem die Korallen leben, hat viel niedrigere 14C-Werte (Matsumoto, 2007; Alves et al., 2018). Der Nachweis von Bomben-14C in Korallen, die in Gewässern wachsen, die keine Bomben-14C-Werte aufweisen (d. h. durch die Belüftung des Ozeans), untermauert, dass schwarze Korallen ihren Kohlenstoff aus von der Oberfläche stammendem POC beziehen (Roark et al., 2009; Sherwood und Edinger, 2009; Williams und Grottoli, 2010; Prouty et al., 2011). Das bedeutet auch, dass wir keinen Tiefenwasserspeichereffekt modellieren müssen, wie es bei den Steinkorallen der Fall ist, die ihren Skelettkohlenstoff aus DIC in den sie umspülenden Zwischengewässern beziehen.
Vergleiche mit neueren neuseeländischen und globalen Altersdaten von Schwarzen Korallen
Die aus diesem Projekt abgeleiteten Radiokohlenstoff-Altersschätzungen und Wachstumsraten ähneln den Ergebnissen einer Reihe anderer Alters- und Wachstumsstudien, die vor kurzem für wichtige Tiefsee-Schwarzkorallen (Antipatharia) in der Region Neuseeland durchgeführt wurden (Tabelle 5). Diese Alters- und Wachstumsschätzungen für B. patula ähneln anderen Korallengattungen wie Leiopathes-, Antipathes- und Antipathella-Kolonien, die bis zu 2600 Jahre leben und in der Größenordnung von 1-143 μm/Jahr wachsen (Hitt et al., 2020). Unsere Ergebnisse sind ein weiterer Beleg dafür, dass neuseeländische Schwarzkorallen zu den am langsamsten wachsenden Meerestieren der Region gehören, wobei einige Arten eine extreme Langlebigkeit aufweisen.

Tabelle 5. Radiokohlenstoff-Altersschätzungsstudien an neuseeländischen schwarzen Korallen (Antipatharia).
Die Ergebnisse von B. patula und anderen schwarzen Korallen, die in Neuseeland gealtert wurden, zeigen keine klaren Trends in den Wachstumsraten oder der Langlebigkeit in Bezug auf die Wassermassen und Tiefen, in denen diese Korallen leben. Die Proben von der Chatham Rise und der Bay of Plenty stammten meist aus recht ähnlichen Wassertiefen zwischen 750 und <1269 m. Die tieferen Exemplare der B. patula von der Chatham Rise stammten aus einer Tiefe zwischen 1120 und 1269 m (Tabellen 1, 5). Die radiale Wachstumsrate unserer Chatham Rise-Proben (NIWA-49468 und NIWA47911) war mit 20-36 μm/Jahr höher als die der Bay of Plenty-Probe (NIWA-85940) mit 11 μm/Jahr. Da für die B. patula-Proben aus diesen beiden Regionen nur drei Wachstumsraten ermittelt werden konnten, können keine Annahmen über die regionalen Unterschiede bei den Wachstumsraten getroffen werden. Das Spektrum der Wachstumsdaten für schwarze Korallen von Chatham Rise zeigt, dass eine Probe von Antipathes sp. mit 91-143 μm/Jahr die schnellste bisher beobachtete Wachstumsrate aufwies, während eine Probe von Leiopathes secunda mit 5-15 μm/Jahr eine der langsamsten war. Die schwarzen Korallen der Bay of Plenty, Bathypathes patula, Leiopathes secunda und Leiopathes sp. weisen mittlere bis geringe radiale Wachstumsraten von 5-40 μm/Jahr auf. Im Gegensatz dazu hat die Flachwasser-Fiordland-Schwarzkoralle Antipathella fiordensis eine relativ hohe radiale Wachstumsrate (50-57 μm/Jahr), aber es ist unklar, ob dies von der Art oder den Umweltbedingungen im Fjord abhängt oder eine Mischung aus beidem ist.
Diese Forschung stimmt auch mit globalen Studien überein, die zeigen, dass Schwarzkorallen zu den am langsamsten wachsenden Tiefseekorallen gehören. Sherwood und Edinger (2009) geben Altersschätzungen für die schwarze Koralle Stauropathes arctica von bis zu 82 Kalenderjahren an. Prouty et al. (2011) verwendeten Radiokohlenstoff, um Altersschätzungen für lebende Kolonien von Leiopathes sp. bis 2100 Jahre zu erstellen. Carreiro-Silva et al. (2013) schätzten das Alter einer Azoren-Kolonie von Leiopathes sp. auf 2320 Jahre. Ein Exemplar von Leiopathes annosahas aus Hawaii hatte das Potenzial, eine Lebensdauer von mehr als 4000 Jahren zu erreichen (Roark et al., 2009).
Diese neuen Alters- und Wachstumsraten für die empfindliche schwarze Tiefseekoralle B. patula zeigen, dass es sich um eine langlebige Art handelt, die in dieser Studie ein Alter von mehr als 385 Jahren erreichen kann und langsame lineare und radiale Wachstumsraten aufweist. Die Ergebnisse deuten auf eine wahrscheinlich geringe Widerstandsfähigkeit und eine langsame Erholungszeit nach Störungen/Entfernungen hin. Diese Forschungsergebnisse werden als Grundlage für künftige Risikobewertungen und Bewirtschaftungsstrategien im neuseeländischen Rahmen für die Bewirtschaftung von Korallen dienen (Freeman und Cryer, 2019). Das langsame Wachstum und die extreme Langlebigkeit machen außerdem deutlich, dass diese Korallen sehr anfällig für unterseeische Störungen wie Fischerei und Bergbau sind und Jahrhunderte brauchen können, um sich von anthropogenen Einflüssen zu erholen (Hitt et al., 2020).
Schlussfolgerung
Das Alter und die Wachstumsrate von Proben der schwarzen Tiefseekoralle B. patula aus zwei Regionen Neuseelands, der Bay of Plenty und dem Chatham Rise, wurden mit zwei verschiedenen Methoden bestimmt: durch Zählen von Wachstumsringen (Sklerochronologie) und durch Radiokohlenstoffdatierung. Für die Zählung der Jahresringe wurden zwei verschiedene Protokolle verwendet, wobei dunkle und helle Wachstumsschichten verwendet wurden. Diese lieferten unterschiedliche Ergebnisse, die auch nicht mit den Radiokohlenstoff-Altersangaben für mehrere dieser Korallen übereinstimmten. Die Radiokohlenstoffalter von B. patula deuten darauf hin, dass diese Korallenart sehr lange lebt, mit einem Alter von über 385 Jahren und langsamen linearen (5,2-9,6 mm/Jahr) und radialen (11,1-35,7 μm/Jahr) Wachstumsraten. Diese Ergebnisse sind mit dem Alter und den Wachstumsraten anderer schwarzer Korallengattungen aus denselben Regionen Neuseelands und weltweit vergleichbar. Alter und Wachstumsraten von Tiefseearten sind entscheidend, um das potenzielle Risiko dieser Organismen für anthropogene Störungen zu bestimmen. Diese Ergebnisse werden für zukünftige Risikobewertungen und Managementstrategien im neuseeländischen Rahmen für das Management von Korallen verwendet.
Datenverfügbarkeitserklärung
Alle für diese Studie generierten Datensätze sind im Artikel/Ergänzungsmaterial enthalten.
Beiträge der Autoren
PM, DT, HB und NH trugen zur Konzeption und zum Design der Studie bei. PM führte die gesamte Probenvorbereitung, die Bildanalyse der Schnitte, die Entwicklung des Dünnschliffprotokolls und die Zählung der Zonen sowie das Mikrofräsen der Radiokohlenstoffproben durch. NH untersuchte die Radiokohlenstoffproben. SF erstellte korrigierte Altersschätzungen aus den Radiokohlenstoff-Rohdaten und schrieb Teile des Manuskripts. PM und DT verfassten den ersten Entwurf des Manuskripts. Alle Autoren trugen zur Überarbeitung des Manuskripts bei.
Finanzierung
Diese Forschungsarbeit wurde aus dem zweiten Jahr des Conservation Services Programme (CSP), Department of Conservation (DOC) Projekt POP2017-07 finanziert, das zum Ziel hat, „eine Methodik zur Bestimmung des Alters und der Wachstumscharakteristika von wichtigen Hochrisikokorallenarten der neuseeländischen Tiefsee (Kaltwasser) zu entwickeln.“ Die Finanzierung der Publikationsgebühr für diese Zeitschrift wurde vom NIWA, Neuseeland, zur Verfügung gestellt.
Interessenkonflikt
Die Autoren erklären, dass die Forschung in Abwesenheit von kommerziellen oder finanziellen Beziehungen durchgeführt wurde, die als potenzieller Interessenkonflikt ausgelegt werden könnten.
Danksagungen
Wir danken der Conservation Services Programme Group, Department of Conservation New Zealand, insbesondere Ian Angus, Kris Ramm, und auch Freydis Hjorvarsdottir (früher CSP, jetzt Ministry of Primary Industries New Zealand) für ihre Unterstützung der Arbeit. Wir danken Helen Neil (NIWA) und Dan Sinclair (VUW) für die Unterstützung bei der Bereitstellung ihrer vorläufigen Daten. Wir danken den NIWA-Mitarbeitern Sadie Mills und Diana Macpherson für ihren Beitrag zur Auswahl der Proben, Owen Anderson für die Erstellung von Abbildung 1, Peter Horn für eine gründliche interne Überprüfung und Rosemary Hurst für hilfreiche Kommentare und Ratschläge zum Manuskript. Wir danken Jeremy Horowitz (James Cook University, Australien) für die Identifizierung der Proben und Dennis Opresko (Emeritus Smithsonian Institute) für Kommentare zur Überarbeitung der Taxonomie der Schwarzen Koralle. Schließlich danken wir den drei Gutachtern, deren detaillierte und konstruktive Bewertung diesem Dokument sehr zugute kam.
Adkins, J. F., Griffin, S., Kashgarian, M., Cheng, H., Druffel, E. R. M., Boyle, E. A., et al. (2002). Radiokohlenstoffdatierung von Tiefseekorallen. Radiocarbon 44, 567-580. doi: 10.1017/s0033822200031921
CrossRef Full Text | Google Scholar
Althaus, F., Williams, A., Schlacher, T. A., Kloser, R. J., Green, M. A., Barker, B. A., et al. (2009). Die Auswirkungen der Grundschleppnetzfischerei auf die Tiefsee-Ökosysteme von Seebergen sind lang anhaltend. Mar. Ecol. Prog. Ser. 397, 279-294. doi: 10.3354/meps08248
CrossRef Full Text | Google Scholar
Alves, E. Q., Macario, K., Ascough, P., and Bronk Ramsey, C. (2018). The worldwide marine radiocarbon reservoir effect: definitions, mechanisms, and prospects. Rev. Geophys. 56, 278-305. doi: 10.1002/2017RG000588
CrossRef Full Text | Google Scholar
Anderson, O., Tracey, D., Bostock, H., Williams, M., and Clark, M. (2014). Refined Habitat Suitability Modelling for Protected Coral Species in the New Zealand EEZ. NIWA-Kundenbericht für das Department of Conservation. WLG2014-69No. 46. Auckland: NIWA.
Google Scholar
Andrews, A. H., Cailliet, G. M., Kerr, L. A., Coale, K. H., Lundstrom, C., and DeVogelaere, A. P. (2005). „Investigations of age and growth for three deep-sea corals from the Davidson Seamount off central California,“ in Cold-Water Corals and Ecosystems, eds A. Freiwald and J. M. Roberts (Berlin: Springer-Verlag), 1021-1038. doi: 10.1007/3-540-27673-4_51
CrossRef Full Text | Google Scholar
Andrews, A. H., Cordes, E. E., Mahoney, M. M., Munk, K., Coale, K. H., Cailliet, G. M., et al. (2002). Alter, Wachstum und radiometrische Altersbestimmung einer habitatbildenden Tiefsee-Gorgonie (Primnoa resedaeformis) aus dem Golf von Alaska. Hydrobiologia 471, 101-110.
Google Scholar
Anthony, K. R. N., and Marshall, P. A. (2009). Coral Reefs and Climate Change. Marine Climate Change Impacts and Adaptation Report Card for Australia 2009. Gland: IUCN.
Google Scholar
Bronk Ramsey, C. (2001). Entwicklung des Radiokohlenstoff-Kalibrierungsprogramms OxCal. Radiocarbon 43, 355-363. doi: 10.1017/s0033822200038212
CrossRef Full Text | Google Scholar
Cairns, S., Gershwin, L., Brook, F., Pugh, P., Dawson, E., Ocanþa, O., et al. (2009). „Phylum cnidaria: corals, medusae, hydroids myxozo-ans,“ in New Zealand Inventory of Biodiversity, Volume 1: Kingdom Animalia, ed. D. P. Gordon (New Zealand: Canterbury University Press), 59-101.
Google Scholar
Carreiro-Silva, M., Andrews, A. H., Braga-Henriques, A., de Matos, V., Porteiro, F. M., and Santos, R. S. (2013). Variabilität der Wachstumsraten der langlebigen schwarzen Koralle Leiopathes sp. von den Azoren. Mar. Ecol. Progr. Ser. 473, 189-199. doi: 10.3354/meps10052
CrossRef Full Text | Google Scholar
Chiswell, S. M., Bostock, H. C., Sutton, P. J. H., and Williams, M. J. M. (2015). Physikalische Ozeanographie der Tiefsee um Neuseeland: ein Überblick, New Zealand. J. Mar. Freshw. Res. 49, 1-32. doi: 10.1080/00288330.2014.992918
CrossRef Full Text | Google Scholar
Clark, M., Bowden, D., Baird, S., and Stewart, R. (2010). Auswirkungen der Fischerei auf die benthische Biodiversität der Seeberge des „Graveyard“-Komplexes, nördlicher Chatham Rise. New Zeal. Aqu. Environ. Biodivers. Rep. 46:40.
Google Scholar
Clark, M., and Rowden, A. (2009). Effects of deepwater trawling on the macro-invertebrate assemblages of seamounts on the Chatham Rise, New Zealand. Deep Sea Res. I 56, 1540-1554. doi: 10.1016/j.dsr.2009.04.015
CrossRef Full Text | Google Scholar
Clark, M., Tracey, D., Anderson, O., and Parker, S. (2014). Pilot Ecological Risk Assessment for Protected Corals. Wellington: National Institute of Water and Atmospheric Research, 32.
Google Scholar
Clark, M. R., Bowden, D. A., Rowden, A. A., and Stewart, R. (2019). Wenig Beweise für die Widerstandsfähigkeit benthischer Gemeinschaften gegenüber der Grundschleppnetzfischerei auf Seebergen nach 15 Jahren. Front. Mar. Sci. 6:63. doi: 10.3389/fmars.2019.00063
CrossRef Full Text | Google Scholar
Coleman, D. C. (1991). Carbon Isotope Techniques. Cambridge, MA: Academic Press.
Google Scholar
Consalvey, M., MacKay, K., and Tracey, D. (2006). Information review for protected deep-sea coral species in the New Zealand region. NIWA Kundenbericht. WLG2006-85. 60. Auckland: NIWA.
Google Scholar
Druffel, E. R. M., Griffin, S., Witter, A., Nelson, E., Southon, J., Kashgarian, M., et al. (1995). Gerardia: Borstenkiefer der Tiefsee? Geochim. Cosmochim. Acta 59, 5031-5036. doi: 10.1016/0016-7037(95)00373-8
CrossRef Full Text | Google Scholar
Druffel, E. R. M., King, L. L., Belastock, R. A., and Buesseller, K. O. (1990). Die Wachstumsrate einer Tiefseekoralle anhand von 210Pb und anderen Isotopen. Geochim. Cosmochim. Acta 54, 1493-1500.
Google Scholar
Fallon, S. J., Fifield, L. K., and Chappell, J. M. (2010). Das nächste Kapitel der Radiokohlenstoffdatierung an der Australian National University: Statusbericht über die einstufige AMS. Nuclear Instr. Methods Phys. Res. Sec. B 268, 898-901. doi: 10.1016/j.nimb.2009.10.059
CrossRef Full Text | Google Scholar
Freeman, D., and Cryer, M. (2019). „Current management measures and threats,“ in The State of Knowledge of Deep-Sea Corals in the New Zealand Region, Chap. 9, eds D. M. Tracey and F. Hjorvarsdottir (Auckland: NIWA Science and Technology Series).
Google Scholar
Gammon, M. J., Tracey, D. M., Marriott, P. M., Cummings, V. J., and Davy, S. K. (2018). Die physiologische Reaktion der Tiefseekoralle Solenosmilia variabilis auf die Ozeanversauerung. PEERJ 6:e5236. doi: 10.7717/peerj.5236
PubMed Abstract | CrossRef Full Text | Google Scholar
Griffin, S., and Druffel, E. M. (1989). Quellen von Kohlenstoff für Tiefseekorallen. Radiocarbon 31, 533-543. doi: 10.1017/s0033822200012121
CrossRef Full Text | Google Scholar
Hitt, N. T., Sinclair, D. J., Fallon, S. J., Neil, H. L., Tracey, D. M., Komugabe-Dixson, A., et al. (2020). Wachstum und Langlebigkeit von neuseeländischen schwarzen Korallen. Deep Sea Res. I 2020:103298. doi: 10.1016/j.dsr.2020.103298
CrossRef Full Text | Google Scholar
Komugabe, A. F., Fallon, S. J., Thresher, R. E., and Eggins, S. M. (2014). Modern Tasman Sea surface reservoir ages from deep-sea black corals. Deep-Sea Res. 99, 207-212. doi: 10.1016/j.dsr2.2013.05.024
CrossRef Full Text | Google Scholar
Komugabe-Dixson, A. F., Fallon, S. J., Eggins, S. M., and Thresher, R. E. (2016). Radiokohlenstoff-Beweise für mittelspäte holozäne Veränderungen in der Zirkulation des südwestlichen Pazifiks. Paleoceanography 31, 971-985. doi: 10.1002/2016PA002929
CrossRef Full Text | Google Scholar
Love, M. S., Yoklavich, M. M., Black, B. A., and Andrews, A. H. (2007). Alter von Kolonien der Schwarzen Koralle (Antipathes dendrochristos), mit Anmerkungen zu den dazugehörigen wirbellosen Arten. Bull. Mar. Sci. 80, 391-400.
Google Scholar
Matsumoto, K. (2007). Radiokarbonbasiertes Zirkulationsalter der Weltmeere. J. Geophys. Res. 112:C09004. doi: 10.1029/2007JC004095
CrossRef Full Text | Google Scholar
Mortensen, P. B., und Buhl-Mortensen, L. (2005). Morphologie und Wachstum der Tiefwassergorgonien Primnoa resedaeformis und Paragorgia arborea. Mar. Biol. 147, 775-788. doi: 10.1007/s00227-005-1604-y
CrossRef Full Text | Google Scholar
Noé, S. U., Lembke-Jene, L., and Chr Dullo, W. (2008). Unterschiedliche Wachstumsraten bei Bambuskorallen: Sklerochronologie und Radiokarbondatierung eines mittelholozänen Tiefsee-Gorgonien-Skeletts (Keratosis sp.: Octocorallia) von Chatham Rise (Neuseeland). Facies 54, 151-166. doi: 10.1007/s10347-007-0129-x
CrossRef Full Text | Google Scholar
Pinkerton, M. H. (2016). Ocean Colour Satellite Observations of Phytoplankton in the New Zealand EEZ, 1997-2016. Ministry for the Environment. Wellington: NIWA.
Google Scholar
Prouty, N. G., Fisher, C. R., Demopoulous, A. W. J., and Druffel, E. R. M. (2016). Wachstumsraten und Alter von Tiefseekorallen, die von der Deepwater Horizon Ölpest betroffen sind. Deep-Sea Res. II 129, 196-212. doi: 10.1016/j.dsr2.2014.10.021
CrossRef Full Text | Google Scholar
Prouty, N. G., Roark, E. B., Buster, N. A., and Ross, S. W. (2011). Wachstumsrate und Altersverteilung von schwarzen Tiefseekorallen im Golf von Mexiko. Mar. Ecol. Prog. Ser. 423, 101-115. doi: 10.3354/meps08953
CrossRef Full Text | Google Scholar
Reimer, P. J., Bard, E., Bayliss, A., Beck, J. W., Blackwell, P. G., Bronk Ramsey, C., et al. (2013). IntCal13 und Marine13 Radiokohlenstoff-Alterskalibrierungskurven 0-50.000 Jahre cal BP. Radiocarbon 55, 1869-1887. doi: 10.2458/azu_js_rc.55.16947
PubMed Abstract | CrossRef Full Text | Google Scholar
Reimer, P. J., and Reimer, R. W. (2001). Eine Datenbank zur Korrektur mariner Lagerstätten und eine Online-Schnittstelle. Radiocarbon 43, 461-463. doi: 10.1017/s0033822200038339
CrossRef Full Text | Google Scholar
Roark, E. B., Guilderson, T. P., Dunbar, R. B., Fallon, S. J., und Mucclarone, D. A. (2009). Extreme Langlebigkeit bei proteinhaltigen Tiefseekorallen. Proc. Natl. Acad. Sci. U.S.A. 106, 5204-5208. doi: 10.1073/pnas.0810875106
PubMed Abstract | CrossRef Full Text | Google Scholar
Roark, E. B., Guilderson, T. P., Dunbar, R. B., and Ingram, B. L. (2006). Radiokohlenstoff-basierte Alter und Wachstumsraten: Hawaiianische Tiefseekorallen. Mar. Ecol. Progr. Ser. 327, 1-37.
Google Scholar
Sánchez, J., and Rowden, A. (2006). „Octocoral diversity on New Zealand Seamounts“, in Proceedings of the Presented at the 10th International Coral Reef Symposium, Okinawa.
Google Scholar
Sherwood, O. A., and Edinger, E. N. (2009). Alter und Wachstumsraten einiger Tiefsee-Gorgonien und antipatharischer Korallen von Neufundland und Labrador. Can. J. Fish. Aquatic Sci. 66, 142-152. doi: 10.1139/f08-195
CrossRef Full Text | Google Scholar
Sherwood, O. A., Scott, D. B., Risk, M. J., and Guilderson, T. P. (2005). Radiokohlenstoff-Beweise für jährliche Wachstumsringe in der Tiefsee-Krakenmuschel Primnoa resedaeformis. Mar. Ecol. Prog. Ser. 301, 129-134. doi: 10.3354/meps301129
CrossRef Full Text | Google Scholar
Sikes, E., Burgess, S., Grandpre, R., and Guilderson, T. (2008). Bewertung moderner Tiefseealter in der neuseeländischen Region anhand von Tiefseekorallen. Deep Sea Res. I 55, 38-49. doi: 10.1016/j.dsr.2007.10.004
CrossRef Full Text | Google Scholar
Stuiver, M., and Polach, H. A. (1977). Discussion reporting of 14C data. Radiocarbon 19, 355-363. doi: 10.1017/S0033822200003672
CrossRef Full Text | Google Scholar
Thresher, R., Rintoul, S. R., Koslow, J. A., Weidman, C., Adkins, J., and Proctor, C. (2004). Ozeanische Belege für den Klimawandel in Südaustralien in den letzten drei Jahrhunderten. Geophys. Res. Lett. 31:l07212.
Google Scholar
Thresher, R. E., MacRae, C. M., Wilson, N. C., und Gurney, R. (2007). Umwelteinflüsse auf die Skelettzusammensetzung von Tiefwassergorgonien (Keratoisis spp.; Isididae). Bull. Mar. Sci. 8, 409-422.
Google Scholar
Tracey, D., Bostock, H., and Shaffer, M. (2018). Ageing Methods for Protected Deep-Sea Corals: A Review and Recommendation for an Ageing Study. DOC Contract 4527 GMC – Age and Growth of coral (POP2017-07). NIWA Client Report WN2018035. Auckland: NIWA, 40.
Google Scholar
Tracey, D., Neil, H., Gordon, D., and O’Shea, S. (2003). Chroniken der Tiefe: Alterung von Tiefseekorallen in neuseeländischen Gewässern. NIWA Water Atmosph. 11, 22-24.
Google Scholar
Tracey, D. M., and Hjorvarsdottir, F. (eds) (2019). „The state of knowledge of deep-sea corals in the new zealand region“, in NIWA Science and Technology Series Number 84, (Auckland: NIWA),140.
Google Scholar
Tracey, D. M., Neil, H., Marriott, P., Andrews, A. H., Cailliet, G. M., and Sanchez, J. A. (2007). Alter und Wachstum von zwei Gattungen von Tiefsee-Bambuskorallen (Familie Isididae) in neuseeländischen Gewässern. Bull. Mar. Sci. 81, 393-408.
Google Scholar
Williams, A., Schlacher, T. A., Rowden, A. A., Althaus, F., Clark, M. R., Bowden, D. A., et al. (2010). Megabenthische Seegraswiesen erholen sich nicht von den Auswirkungen der Schleppnetzfischerei. Mar. Ecol. 31, 183-199. doi: 10.1111/j.1439-0485.2010.00385.x
CrossRef Full Text | Google Scholar
Williams, B., and Grottoli, A. G. (2010). Variabilität der stabilen Stickstoff- und Kohlenstoffisotope (δ15N und δ13C) in flachen tropischen pazifischen Weichkorallen und schwarzen Korallentaxa und Implikationen für paläozeanographische Rekonstruktionen. Geochim. Cosmochim. Acta 74, 5280-5288. doi: 10.1016/j.gca.2010.06.026
CrossRef Full Text | Google Scholar
Williams, B., Risk, M. J., Ross, S. W., and Sulak, K. J. (2007). Stabile Isotopendaten von Tiefsee-Antipathariern: 400-Jahres-Aufzeichnungen von der südöstlichen Küste der Vereinigten Staaten von Amerika. Bull. Mar. Sci. 81, 437-447.
Google Scholar
Leave a Reply