Frontiers in Marine Science
Úvod
Hlubokomořští koráli jsou velmi rozmanitou skupinou mořských organismů a oblast Nového Zélandu je horkým místem pro hlubokomořské korály (Sánchez a Rowden, 2006; Cairns a kol., 2009). Několik studií zkoumalo vliv lovu vlečnými sítěmi při dně na společenstva hlubokomořských makrobezobratlých (včetně korálů) v oblasti jižního Pacifiku (Althaus a kol., 2009; Clark a Rowden, 2009; Clark a kol., 2010, 2019; Williams a kol., 2010; Anderson a kol., 2014). Ty prokázaly znatelné dopady lovu vlečnými sítěmi při dně na pomalu rostoucí a křehké hlubokomořské korálové kolonie a společenstva s malou nebo žádnou schopností obnovy. Očekává se, že environmentální dopady předpovídané v důsledku změny klimatu, jako je oteplování oceánů a okyselování oceánů, rovněž ohrozí zdraví hlubokomořských korálů, přičemž některá stanoviště se stanou příliš teplými na to, aby se jim dařilo Anthony a Marshall (2009), nebo se sníží pH a obsah uhličitanových iontů, což bude mít pravděpodobně dopad na hlubokomořské korály s karbonátovou kostrou (Gammon et al., 2018). Pro lepší informovanost o budoucích hodnoceních rizik a řízení mořských zdrojů je pochopení stáří a růstu hlubokomořských korálů klíčové pro určení doby regenerace korálů a jejich obnovitelnosti po antropogenních narušeních nebo jiných přírodních škodách.
V oblasti Nového Zélandu bylo provedeno značné množství prací týkajících se taxonomie hlubokomořských korálů, rozšíření různých druhů a jejich environmentálních preferencí, jakož i významu hlubokomořských korálů pro bentické ekosystémy. Nedávno byl publikován přehled našich znalostí a vědomostí o hlubinných korálech v oblasti Nového Zélandu (Tracey a Hjorvarsdottir, 2019). Tento přehled také nastínil hlavní mezery v našem poznání hlubokomořských korálů, včetně omezených údajů o stáří a růstu mnoha rodů a druhů. V této studii hodnotíme věk a rychlost růstu chráněného hlubokomořského černého korálu Antipatharian Bathypathes patula. Věk a rychlost růstu tohoto korálu nebyly v této oblasti dosud studovány. Výběr tohoto druhu byl založen na seznamu priorit hodnocení rizik (Clark et al., 2014) a přehledu literatury (Tracey et al., 2018) spolu s dostupností vzorků (lokalita a celkový počet) a dalším doplňujícím výzkumem černých korálů prováděným na Novém Zélandu (Hitt et al., 2020).
Tracey et al. (2018) nastínili současné metody, které se využívají ke stanovení stáří hlubokomořských druhů korálů. Byly diskutovány výhody a nevýhody jednotlivých metod. Hlavními metodami, které byly úspěšně použity k měření stáří a růstu hlubokomořských korálů, jsou (1) přímé pozorování, např. měření in situ nebo experimenty v akváriu s lineárním růstem nebo povrchovým rozšířením; rychlost přírůstku polypů; odhad rychlosti kalcifikace (např, pomocí techniky vztlakové váhy), (2) výčet kosterních růstových zón nebo pásů (sklerochronologie) a (3) radiometrické analýzy.
Přímá pozorování a měření in situ nebo v akváriu je u hlubokomořských korálů velmi obtížné provádět, protože je obtížné získat živé životaschopné exempláře a udržet je při životě, protože jejich růst trvá dlouho (Gammon et al., 2018; Tracey et al., 2018). Vyčíslování růstových zón, sklerochronologie, je ideální pro hlubokomořské druhy korálů, které mají vysoký kontrast mezi zónovými růstovými vzorci pozorovanými v kosterních řezech. Je však nezbytné, aby byla použita nezávislá validační metoda k ověření chronologické platnosti počtů pozorovaných růstových zón (Andrews et al., 2002). Metoda získávání počtů viditelných růstových zón v kosterní struktuře byla úspěšně použita ke stanovení údajů o stáří a rychlosti růstu gorgonských oktokorálů včetně 400 let starého bambusového korálu Keratoisis sp., 100 let starých mořských korálů Primnoa spp. a 60 let starých bambusových korálů Lepidisis sp. (Thresher et al., 2004, 2007; Mortensen a Buhl-Mortensen, 2005; Sherwood et al., 2005; Tracey et al., 2007; Sherwood a Edinger, 2009). Stáří černých korálů bylo také úspěšně určeno pomocí růstových zón nebo počtů, přičemž stáří korálů Antipathes dendrochristos a Leiopathes glaberrima se pohybuje od 150 do 480 let (Love et al., 2007; Williams et al., 2007). Ve studii černého korálu Stauropathes arctica byly získány zónové počty 55-58 let (Sherwood a Edinger, 2009), přičemž autoři poznamenávají, že radiokarbonové datování omezilo toto stáří na 55 a 82 let. Rychlost růstu černých korálů pomocí výčtu růstových zón ukazuje nízký radiální růst od 8 do 140 μm/rok (Love et al., 2007; Prouty et al., 2011).
Metoda výčtu růstových zón má potenciální omezení (viz shrnutí v Tracey et al., 2018). Mnoho korálů nevykazuje jasnou pravidelnou zónovou strukturu v celém svém skeletu nebo může být jasná zónová struktura patrná pouze v částech skeletu. I když je zónová struktura přítomna, může být proměnlivá nebo vykazovat různé možnosti interpretace. Například bazální a hrotové počty bambusových oktokorálů Keratoisis sp. a Lepidisis sp. byly vyšší než stáří odhadnuté radiometrickou metodou olova 210 (210Pb) (Tracey et al., 2007). Byla vyslovena hypotéza, že zóny pozorované světelným mikroskopem mají dvouletou periodicitu a že zóny pozorované SEM v uzlovém spoji mohou představovat environmentální událost, např. měsíční periodicitu. Lunární cykly mohou být spojeny se silnými přílivovými a odlivovými událostmi, které by mohly zvýšit dostupnost potravy pro přisedlé organismy živící se filtrací, což by vedlo k pulzům somatického růstu. Stáří gorgonských bublinatých korálů (např. Paragorgia spp.) a červených drahokamových korálů (např. Corallium sp.) nebylo úspěšně určeno pomocí počítání zón kvůli složitým nebo špatně definovaným vzorům pásů (Griffin a Druffel, 1989; Andrews et al.,
Nejběžnější radiometrickou metodou používanou u hlubokomořských korálů je datování pomocí radiouhlíkového izotopu (14C) (Druffel et al., 1995; Sherwood et al., 2005; Tracey et al., 2018). Tato metoda se opírá o skutečnost, že velmi malé množství přírodního uhlíku v atmosféře je radioaktivní ve formě 14C a je zabudováno do kostry korálů při tvorbě jejich uhličitanu vápenatého nebo bílkovin a chitinu (Adkins et al., 2002; Tracey et al., 2003; Consalvey et al., 2006). Protože poločas rozpadu radiouhlíku je, jak známo, 5730 let, lze tuto metodu, byť drahou, použít ke spolehlivému zestárnutí vzorků starých méně než 50 000 let; za touto hranicí se aktivita 14C stává příliš nízkou na to, aby ji bylo možné zjistit (Coleman, 1991). V 50.-60. letech 20. století došlo k rychlému nárůstu obsahu 14C v atmosféře v důsledku testování jaderných zařízení; tento „bombový uhlík“ lze rovněž použít jako nástroj pro kalibraci stáří, který poskytuje referenční bod pro novější vzorky (Coleman, 1991; Druffel et al., 1995; Tracey et al., 2003; Roark et al., 2009; Sherwood a Edinger, 2009).
Radiouhlíkové datování bylo úspěšně použito u několika druhů korálů, včetně kamenných korálů, černých korálů a gorgonských oktokorálů (Druffel et al., 1990; Roark et al., 2006; Carreiro-Silva et al., 2013; Prouty et al., 2016; Hitt et al., 2020). Tato metoda má však také svá omezení. V důsledku oceánské cirkulace je radiouhlíkové stáří vody v oceánech výrazně starší než v atmosféře, tzv. mořské zásobníkové stáří (Tracey et al., 2003; Consalvey et al., 2006; Roark et al., 2006; Noé et al., 2008). Chyby v zásobníkovém stáří mohou mít významný vliv na odhady stáří korálů. Korekce stáří rezervoáru vyžaduje znalost okolních vod a předpokládá konstantní poměry radiouhlíku ve vodě; tyto hladiny však mohou být změněny mícháním v oceánu, lokálními přívalovými jevy a produktivními květy, což vše může změnit přeměny uhlíkového cyklu a složení 14C v kostře korálů (Tracey et al.),
Na základě přehledu Tracey et al. (2018) jsme se rozhodli použít dvě metody stárnutí B. patula; počty kosterních růstových pásů a radiometrickou metodu radiouhlíkového datování (14C). Použití těchto metod umožňuje srovnání s dalšími předchozími studiemi stárnutí korálů na Novém Zélandu i ve světě. Je uveden výběr vzorků, metody mikrofrézování materiálu a řezání a interpretace počítání pásů a radiouhlíkového datování. Jsou uvedeny věkové a růstové charakteristiky černého korálu B. patula a jedné kolonie kolonie Bathypathes cf. conferta (původně předpokládané jako B. patula).
Materiál a metody
Výběr vzorků
Vzorky použité v této studii pocházely ze zachovaných exemplářů uložených v novozélandském Národním ústavu pro výzkum vody a atmosféry (NIWA), Národní sbírce bezobratlých (NIC).
Vzorky byly vybrány ze dvou oblastí: Bay of Plenty (severovýchodní část Severního ostrova Nového Zélandu) a Chatham Rise (východní část Nového Zélandu), aby bylo možné porovnat výsledky a posoudit variabilitu v rychlosti růstu mezi různými oblastmi a také s dalšími rody hlubokomořských černých korálů, konkrétně Antipathella, Leiopathes a Antipathes, ze stejných oblastí (Hitt et al..), 2020).
Oceánografické proudy ovlivňují hlubokomořské korály tím, že zajišťují stálý přísun potravy a rozptyl larev. V důsledku toho tyto proudy ovlivňují také dostupnost kořisti a rozšíření hlubokomořských korálů. Hlubokomořské korály jsou sice nejvíce ovlivňovány spodními vodními proudy, které nad nimi proudí a ovlivňují distribuci sedimentů, ale jsou také ovlivňovány povrchovými vodními proudy, které ovlivňují primární produktivitu, a tím i nabídku potravy, která padá k hlubokomořským korálům na mořském dně. Pevnina Nového Zélandu se nachází na rozhraní teplých subtropických povrchových vod chudých na živiny, které proudí ze severu, a chladných subantarktických povrchových vod bohatých na živiny, které proudí z jihu. Současné znalosti o vodních masách a oceánských proudech v oblasti na povrchu, v termoklině, ve středních vodách a v hlubokých vodách nedávno zhodnotili Chiswell et al. (2015). Oceánografické podmínky pro korály z této studie jsou následující:
Plný záliv je omýván teplými subtropickými povrchovými vodami s nízkým obsahem živin, které pocházejí z Východoaustralského proudu a přitékají od západu a tvoří Východoaucklandský proud. V oblasti Chatham Rise se setkávají teplé subtropické vody s nízkým obsahem živin (STW) a chladné subantarktické vody s vysokým obsahem živin (SAW), které přicházejí z Jižního oceánu, je to subtropická fronta (STF). Míchání vodních mas STW a SAW na STF má za následek, že Chatham Rise je zónou velmi vysoké primární produktivity, která poskytuje významný zdroj potravy, a tedy i uhlíku pro tyto korály. koncentrace chlorofylu a se v Chatham Rise a Bay of Plenty pohybují v rozmezí 0,3 a 0,48 mg/m3, zatímco v centrálním Pacifiku je to 0,03-0,3 mg/m3 (Pinkerton, 2016). V hloubce 500-1500 m do této oblasti proudí ze severu a jihu chladné (3,5-10 °C), málo slané (34,4-34,6) a s vysokým obsahem kyslíku (200-250 μmol/kg) a živin (dusičnany 15-36 μmol/kg) antarktické intermediální vody (AAIW) (Chiswell et al., 2015).
Korály z Chatham Rise byly vybrány, protože z této oblasti existují moderní údaje o stáří zásobníků 14C (Sikes et al., 2008). Znalost místního zásobního stáří 14C umožňuje přesnější odhad radiouhlíkového stáří korálů.
Vzorky byly vybrány z existujících vzorků nasbíraných rybářstvím Nového Zélandu (FNZ), pozorovateli Ministerstva pro primární průmysl (MPI) a výzkumníky NIWA a jsou uloženy v (NIC). Při rozhodování o výběru místa kolonie a počtu vzorků pomohl graf údajů o rozšíření B. patula (obr. 1). Dva exempláře ze zálivu Bay of Plenty (NIWA24190 a NIWA85940) byly vybrány pro radiokarbonové datování a tenké řezy pro počty zón. Zbytek vzorků byl vybrán z oblasti Chatham Rise.
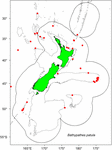
Obrázek 1. Místa, kde byly odebrány vzorky Bathypathes patula v oblasti Nového Zélandu. Červené tečky označují jednotlivá zdrojová místa vzorků korálů B. patula uchovávaných v Národní sbírce bezobratlých Nového Zélandu (NIC). Šedé čáry označují obrys hloubky vody 200 m. Černá čára označuje rozsah výlučné ekonomické zóny Nového Zélandu (EEZ).
Na konci roku 2018 přezkoumal hostující specialista na taxonomii černých korálů (Jeremy Horowitz, James Cook University) identifikace několika vzorků v NIC. Během tohoto procesu byla revidována identifikace jedné z kolonií černých korálů vybraných pro tento výzkum odhadu stáří a předpokládaných jako Bathypathes patula. Černý korál NIWA24190 byl nyní identifikován jako Bathypathes cf. conferta. Vzorek byl v celé této práci označován jako Bathypathes cf. conferta (obr. 2), což je nejpravděpodobnější správná identifikace. Další taxonomické přezkoumání tohoto vzorku probíhá (Opresko pers comm). Mapa rozšíření (obr. 1) může obsahovat další exempláře B. cf. conferta.

Figure 2. Bathypathes cf. conferta. NIWA24190. Vzorek ukazující místo, kde byly odebrány vzorky pro analýzu. (A) je rostoucí špička, (B) bazální řez. Vzorek ze zálivu Bay of Plenty, Nový Zéland. Scale bar is a 35 cm ruler.
Příprava materiálu
Po určení vhodných vzorků byly z rostoucích špiček a bazální části hlavního stonku korálů vyříznuty malé (menší než 10 mm) úseky. Vzhledem k tomu, že mnoho vzorků bylo během sběru zlomeno, byly odebrány další úseky podél hlavního stonku, aby bylo možné vytvořit spolehlivé odhady lineární rychlosti růstu (obr. 2-5). Vzorky z rostoucích špiček byly poté rozděleny na dva fragmenty, jeden pro radiokarbonové datování, druhý pro přípravu tenkých řezů k získání počtu zón.

Obrázek 3. Bathypathes patula NIWA85940. Vzorek ukazující, kde byly odebrány vzorky pro analýzu. (A) je rostoucí špička, (C) bazální řez a (B) je mezilehlé místo odběru vzorků, protože exemplář byl při odběru zlomen a nad tímto místem mohou chybět neznámé délky hlavního kmene větve. Vzorek ze zálivu Bay of Plenty, Nový Zéland. Měřítko je 35cm pravítko.

Obrázek 4. Bathypathes patula NIWA49468. Vzorek ukazující místo, kde byly odebrány vzorky pro analýzu. (A) je rostoucí špička, (D) bazální řez a (B,C) jsou mezilehlá místa odběru vzorků, protože exemplář byl při odběru zlomený a mezi těmito dvěma místy může chybět neznámá délka hlavního kmene větve. Vzorek z Chatham Rise, Nový Zéland. Měřítko je 35cm pravítko.

Obrázek 5. Bathypathes patula NIWA47911. Vzorek ukazující, kde byly odebrány vzorky pro analýzu. (A) je rostoucí špička, (F) bazální řez a (B-E) jsou místa mezi vzorky odebranými za účelem hledání signálu radiouhlíkové bomby. Vzorek z Chatham Rise, Nový Zéland. Scale bar is a 35 cm ruler.
Mikrofrézování materiálu
Z předchozích prací bylo zjištěno, že řezy černých korálů jsou náchylné k nasávání pryskyřice skrz jejich porézní matrici kapilárním působením, a proto řezy v této studii nemohly být spojeny pryskyřicí se základní deskou pro frézování. Místo toho bylo vyrobeno sklíčidlo, které drží řez během frézování, čímž se eliminuje riziko kontaminace radiouhlíkových vzorků pryskyřicí.
Práškované radiouhlíkové vzorky byly extrahovány z okraje a jádra bazálních, středových a špičkových řezů pomocí mikrofrézy New WaveTM s frézou Brassler H2.11.006 o průměru 0,5 mm v NIWA. Okrajové vzorky zahrnovaly materiál z vnějších 250 mikronů řezu, vzorky z jádra byly získány z materiálu v okruhu 750 mikronů od primordia. Snažili jsme se získat alespoň 1,5 mg materiálu z každého vzorku. Vzorky byly následně zváženy a analyzovány na radiouhlík v radiouhlíkové laboratoři Australské národní univerzity (ANU).
Metoda tenkých řezů a protokoly počítání zón
Řez stonkem korálu byl zalit do čiré dvousložkové epoxidové pryskyřice a rozřezán diamantovou pilkou v NIWA. Řezy byly z jedné strany vyleštěny a upevněny leštěnou stranou dolů na skleněné mikroskopické sklíčko, poté byly broušeny a leštěny, dokud neměly optimální tloušťku pro zobrazení struktury růstové zóny pomocí procházejícího světla. Optimální tloušťka řezů tohoto druhu byla přibližně 250-300 μm a odpovídá standardnímu postupu přípravy tenkých řezů (Andrews et al., 2002; Mortensen a Buhl-Mortensen, 2005; Tracey et al., 2007). Počítání periodických růstových pásů nebo přírůstků, u nichž se předpokládá, že se tvoří každoročně, probíhalo po přípravě tenkých řezů v NIWA.
Pro vytvoření počtu zón byly preparáty tenkých řezů hlavních větví prohlíženy pod složeným mikroskopem s procházejícím světlem. Jako pomůcka pro interpretaci zón byly řezy prohlédnuty také pomocí ultrafialového světla podle metodiky Sherwooda a Edingera (2009). Zde byly počty zón provedeny za použití procházejícího světla jasného pole a ultrafialové osvětlení bylo užitečnou pomůckou, která pomohla blíže definovat pozorovanou strukturu zón.
Radiouhlíková metoda datování
Před analýzami byly vzorky očištěny (např. vyluhovány kyselinou) od černých krust a endolitické aktivity, aby se odstranil případný mladší kontaminant 14C, který by mohl změnit výsledky (Adkins et al., 2002). Práškové vzorky korálů byly ošetřeny <1 ml 0,1 M HCL. Kyselina byla poté odpipetována a vzorky byly třikrát promyty miliQ vodou. Vzorek byl poté zmrazen suchým ledem a lyofilizován. Přibližně 1 mg vyčištěného vzorku korálu byl navážen do křemenné zkumavky o průměru 6 mm, bylo přidáno ∼60 mg CuO a stříbrný kalíšek. Zkumavky byly evakuovány na <3e-3 torrů (<0,4 Pa) a uzavřeny hořákem. Poté byly zkumavky vypalovány při teplotě 900 °C po dobu 6 hodin za vzniku CO2. Vzniklý CO2 byl vyčištěn a převeden na grafit za přítomnosti vodíku s poháněným Fe jako katalyzátorem. Grafit byl poté změřen na jednostupňovém urychlovacím hmotnostním spektrometru ANU (Fallon et al., 2010). Všechna data byla korigována pomocí on-line AMS δ13C, normalizována na kyselinu šťavelovou I a pozadí odečteno pomocí uhlí bez 14C zpracovaného stejným způsobem jako vzorky korálů. Data jsou prezentována podle doporučení Stuivera a Polacha (1977).
Výsledky
Tenké řezy pro zónové počty
Zónové strukturní vzorce pozorované na kosterních řezech těchto druhů jsou velmi složité a lze je interpretovat mnoha způsoby. Pro počty radiálního růstu byly připravené řezy pečlivě pozorovány a osa, která nejlépe vykazovala jasnou zónovou strukturu od primordia k vnějšímu okraji, byla poté použita pro získání počtů zón. Radiální růst větví u B. patula byl přiměřeně pravidelný, nedošlo k výrazné odchylce v ukládání zón tvořících silnější zóny na některé konkrétní straně větve, což bylo dříve pozorováno u jiných druhů černých korálů. Poloměry použité pro výpočet rychlosti růstu byly průměrem minimálních a maximálních poloměrů pozorovaných v připravených řezech, což znamená, že uváděná rychlost radiálního růstu bude průměrnou rychlostí radiálního růstu korálu.
Na kostře korálu se při pozorování pod menší silou (10-20×) objevuje počáteční hrubý pásový vzor do značné míry definovaný střídáním tmavších a světlejších zón (obr. 6). Při podrobnějším zkoumání při větším výkonu (100-200×) je patrný i poměrně pravidelný jemný vzor proužkování (obr. 7). Zde jsme použili dva protokoly čtení – počítali jsme jak hrubě vytvořené široké zóny, tak tenké jemně vytvořené zóny (tabulka 1). Všechny zde prezentované počty zón byly provedeny jedním čtecím zařízením (Marriott). Během procesu vývoje protokolů interpretace zón bylo provedeno mnoho odečtů s použitím různých iterací vyvíjených protokolů. Počty zón ze dvou zde prezentovaných protokolů byly vždy odvozeny z vícenásobného počítání napříč řezem.

Obr. 6. Bazální řez korálem NIWA47911 ukazující interpretaci hrubého barevného páskování pozorované zónové struktury. Vzorek vykazuje 54 zón označených červenými tečkami. Vložený čárkovaný obdélník ukazuje centrální část bazálního řezu zobrazeného na obrázku 7.

Obrázek 7. Centrální část bazálního řezu korálu NIWA47911 zobrazující protokol o interpretaci pozorované zónové struktury v jemném měřítku. Vzorek vykazuje 42 zón označených černými tečkami v této vnitřní oblasti řezu; celý řez vykazoval 211 zón.

Tabulka 1. Počty zón z tenkých řezů.
U nejmladší části studovaných vzorků, v oblasti špičky nebo v blízkosti špičky, se počty pohybovaly od 4 do 7 (počty hrubých zón) a od 10 do 19 (počty jemných zón). Ve střední oblasti hlavní větve se počty pohybovaly od 55 do 60 (počty hrubých zón) a od 370 do 374 (počty jemných zón). V základní oblasti kolonie se počty pohybovaly od 37 do 193 (počty v hrubé zóně) a od 78 do 481 (počty v jemné zóně). Složitost zónových vzorců zdůraznila potřebu validace.
Radiouhlíková analýza
Mleté vzorky uvedené v tabulce 2 byly zaslány na ANU k radiouhlíkovému datování. Výsledky ze 17 vzorků korálů jsou uvedeny v radiouhlíkových výsledcích (tabulka 3). Tři vzorky (vzorky 47911-F2; 24190-B1; 85940-B1) v laboratoři selhaly a nezbyl dostatečný materiál pro pokus o duplikát. Radiouhlíkové výsledky ze zbývajících korálů byly kalibrovány na kalendářní stáří pomocí programu OxCal4.3 (Bronk Ramsey, 2001), křivky Marine13 a novozélandského místního deltaR (radiocarbon reservoir offset) -18 ± 36 (Reimer a Reimer, 2001). Rozložení věkových intervalů pro vzorky korálů z doby před bombardováním je uvedeno v tabulce 3. Všechny vzorky korálů použité v této studii byly živé při odlovu, takže všechny uvedené odhady stáří jsou věkem při odlovu, nikoli délkou života vzorku. Předpokládaná délka života tohoto druhu bude přesahovat zde uvedené odhady věku. Korál 85940 byl přibližně 380 ± 50 let starý, korál 49468 přibližně 310 ± 70 let starý a korál 47911 přibližně 120 ± 50 let starý (tabulka 3). Všechny vzorky vnějších okrajů a špiček korálů obsahovaly radiouhlíkové hodnoty F14C >1 (tab. 3), s výjimkou korálů 85940-B2 a 85940-C2, což naznačuje, že tyto korály byly v době sběru živé a obsahovaly uhlík z povrchové vody. Koráli 85940-B2 a 85940-C2 měli na vnějším okraji radiouhlíkové hodnoty F14C <1 (tab. 3), což naznačuje, že část kostry aktivně nepřidávala materiál. To bylo pozorováno již dříve, kdy bazální část korálové kolonie přestala růst, avšak horní část pokračovala v růstu (Komugabe-Dixson et al., 2016).
Tabulka 2. Vzorky odebrané pro radiouhlíkové datování.

Tabulka 3. Výsledky radiouhlíkového datování.
Odhady rychlosti růstu
Odhady rychlosti růstu byly založeny na výsledcích radiouhlíkového datování, protože počítání zón bylo problematické a ukázalo se jako neověřené, takže nemohlo být použito k vytvoření spolehlivých odhadů rychlosti růstu (další diskuse o počítání zón viz oddíl „Bathypathes patula Age and Growth Estimates“). Rychlost růstu byla odvozena z fyzických vzdáleností mezi místy odběru vzorků a z kalibrovaného radiouhlíkového stáří na příslušném místě odběru. Vzhledem k tomu, že některé vzorky během radiouhlíkové analýzy selhaly, bylo možné vytvořit pouze omezený soubor odhadů rychlosti růstu. Radiální rychlost růstu, tedy rychlost, kterou se větev kolonie v průběhu času radiálně zahušťuje, byla odhadnuta vydělením kalibrovaných kalendářních let růstu mezi jádrem a okrajem místa odběru vzorků průměrným poloměrem úseku větve. Radiální růst se pohyboval od 11,1 do 35,7 μm/rok. Rychlost lineárního růstu, tedy rychlost, kterou se větev kolonie v průběhu času podélně prodlužuje, byla odhadnuta vydělením rozdílu mezi kalibrovanými kalendářními roky vzorků jádra lineární vzdáleností mezi dvěma místy odběru vzorků. Byly získány dvě lineární rychlosti růstu, 5,2 a 9,6 mm/rok (tabulka 4).

Tabulka 4. Odhady rychlosti růstu Bathypathes patula.
Diskuse
Odhady věku a růstu Bathypathes patula
Radiouhlíkové výsledky této práce ukazují, že B. patula je dlouhověký druh, jehož stáří přesahuje 385 let. Z údajů o radiouhlíkovém stáří byla získána pomalá lineární (5,2-9,6 mm/rok) a radiální (11,1-35,7 μm/rok) rychlost růstu.
Žádný z protokolů o počtu zón spolehlivě nekoreloval s nezávislými odhady radiouhlíkového stáří. Protokol počítání zón v jemném měřítku vykazoval nejtěsnější korelaci s údaji o stáří 14C, což poskytlo určitou celkovou podporu pro dlouhověkost při porovnání počítání zón s daty 14C. Odhady stáří mezi údaji o radiouhlíkovém stáří a údaji o počítání zón v jemném měřítku běžně vykazovaly rozdíl více než sto let v úsecích větví s kalibrovaným radiouhlíkovým stářím 112-385 let (tab. 1, 3). Rozdíly v odhadovaných stářích ze srovnatelných vzorků pomocí obou metod byly záporné a kladné; při použití zónových počtů nedošlo k podhodnocení nebo nadhodnocení stáří o poměrnou část (tab. 1). To znamená, že pozorovaná zónová struktura nevykazuje roční ani jiný pravidelný chronologický vzorec periodicity a naše protokoly počítání zón nelze použít ke spolehlivému vytváření odhadů stáří B. patula.
Z výsledků okrajového radiouhlíku ze vzorku NIWA-85940 vyplývá, že bazální část (C2) hlavního kmene přestala ukládat kosterní materiál na svém vnějším povrchu 293 let před sklizní a střední část (B2) přestala ukládat kosterní materiál 88 let před sklizní. A to navzdory zjištění, že tyto úseky větvového kmene bezprostředně sousedily s postranními jehlicemi s živými polypy. Optický vzhled těchto úseků nenasvědčoval tomu, že by se na jejich vnějším okraji přestala ukládat zóna. Povrch korálové větve se stále jevil jako relativně nedotčený, nevykazoval žádné známky povrchové eroze nebo změny barvy, jak by se dalo očekávat u korálové větve, která již nepřibírá nový kosterní materiál na svůj vnější povrch. To naznačuje, že pokud stonky větví mohou přestat přibývat kosterní vrstvy, zatímco konce větví pokračují v růstu, pak protokoly metody zónového počítání nemohou být spolehlivě použity pro vytváření odhadů stáří z bazálních řezů pro tento druh korálů.
U exempláře NIWA47911 bylo odebráno pět vzorků z rostoucího konce hlavní osové větve směrem k bázi korálu. Tyto vzorky byly od sebe vzdáleny 25 mm. Jednalo se o pokus lokalizovat výskyt radiouhlíkové bomby v kostře korálů. To by znamenalo, že lineární rychlost růstu by mohla být přesně vyhodnocena pro novější rostoucí špičky větví kolonie. Pokud by bylo možné přesně lokalizovat nárůst radiouhlíku bomby, pak by to také poskytlo přesný spojovací bod pro stanovení korekce 14C, která by se použila na vzorky radiouhlíku před bombardováním této kolonie a na další kolonie ze stejné vodní masy. Bohužel se ukázalo, že všechny tyto vzorky byly odebrány po bombardování, takže tento proces nemohl být dokončen. Budoucí odběr vzorků dále zpět po tomto kmeni větve by byl užitečný pro stanovení tohoto spojovacího bodu a současné lineární rychlosti růstu. V naší práci byla naměřena celková lineární rychlost růstu 5,2-9,6 mm/rok, za předpokladu nejrychlejší rychlosti růstu by bylo třeba odebrat vzorky 700 mm zpět po hlavním větvovém stonku od rostoucí špičky, aby bylo možné s jistotou pozorovat nárůst radiokarbonu bomby v kostře.
Výzkum Love et al. (2007) u černého korálu druhu Antipathes dendrochristos rovněž uvádí dvě interpretace zónové struktury podobné zde popsaným hrubým a jemným protokolům. Tito autoři však zjistili, že hrubší barevná pásková struktura koreluje s jejich výsledky radiouhlíku a 210Pb, které ukazují na roční páskování (Love et al., 2007).
Radiouhlíková měření z této práce vykazovala F14C >1 ve vzorcích z rostoucích špiček větví a povrchu hlavní větve, což naznačuje, že tyto vzorky vykazovaly moderní postbombové úrovně radiouhlíku. Přítomnost postbombového uhlíku pod smíšenou vrstvou oceánu naznačuje, že černí koráli B. patula využívají nedávno exportovaný POC z povrchové vody. V těchto hloubkách je POC jediným zdrojem uhlíku označeným bombovým 14C. Rozpuštěný anorganický uhlík (DIC) okolní vody, ve které koráli žijí, má mnohem nižší hodnoty 14C (Matsumoto, 2007; Alves et al., 2018). Detekce bomby 14C u korálů, které rostou ve vodách, v nichž se bomba 14C nevyskytuje (tj. díky oceánské ventilaci), posiluje domněnku, že černí koráli získávají uhlík z POC pocházejícího z povrchu (Roark et al., 2009; Sherwood a Edinger, 2009; Williams a Grottoli, 2010; Prouty et al., 2011). To také znamená, že nemusíme modelovat efekt hlubokovodního rezervoáru, jako je tomu v případě kamenných korálů, kteří svůj kosterní uhlík získávají z DIC v mezihladinách, které je omývají.
Srovnání s nedávnými novozélandskými a celosvětovými údaji o stáří černých korálů
Radiouhlíkové odhady stáří a rychlosti růstu získané v rámci tohoto projektu jsou podobné výsledkům řady dalších studií stáří a růstu, které byly nedávno provedeny u klíčových hlubokomořských černých korálů (Antipatharia) v oblasti Nového Zélandu (tabulka 5). Tyto odhady věku a růstu pro B. patula jsou podobné jiným rodům korálů, jako jsou kolonie Leiopathes, Antipathes a Antipathella, které se dožívají až 2600 let a rostou řádově 1-143 μm/rok (Hitt et al., 2020). Naše výsledky dále potvrzují, že novozélandští černí koráli patří k nejpomaleji rostoucím mořským živočichům v regionu, přičemž některé druhy vykazují extrémní délku života.

Tabulka 5. Studie odhadu radiouhlíkového stáří novozélandských černých korálů (Antipatharia).
Výsledky z B. patula a dalších černých korálů, které byly stárnuty na Novém Zélandu, neukazují jasné trendy v rychlosti růstu nebo dlouhověkosti s ohledem na vodní masy a hloubky, ve kterých se tyto korály vyskytují. Vzorky z Chatham Rise a Bay of Plenty většinou pocházely z poměrně podobných hloubek, od 750 do <1269 m. Hlouběji získaný vzorek B. patula z Chatham Rise pocházel z hloubky někde mezi 1120 a 1269 m (tab. 1, 5). Radiální rychlost růstu našich vzorků z Chatham Rise (NIWA-49468 a NIWA47911) 20-36 μm/rok byla vyšší než u vzorku ze zálivu Bay of Plenty (NIWA-85940) 11 μm/rok. Vzhledem k tomu, že se podařilo získat pouze tři růstové rychlosti pro vzorky B. patula z těchto dvou oblastí, nelze učinit žádné předpoklady o regionálních rozdílech v růstových rychlostech. Spektrum údajů o rychlosti růstu černých korálů z Chatham Rise ukazuje, že vzorek Antipathes sp. měl dosud nejrychlejší pozorovanou rychlost růstu, 91-143 μm/rok, stejně jako jednu z nejpomalejších, 5-15 μm/rok u vzorku Leiopathes secunda. Vzorky černých korálů z Bay of Plenty Bathypathes patula, Leiopathes secunda a Leiopathes sp. vykazují střední až nízkou radiální rychlost růstu, 5-40 μm/rok. Naproti tomu mělkovodní fiordlandští černí koráli Antipathella fiordensis mají poměrně vysokou radiální rychlost růstu (50-57 μm/rok), ale není jasné, zda je to dáno druhem nebo podmínkami prostředí Fiordu, případně kombinací obojího.
Tento výzkum se také shoduje s celosvětovými studiemi, které ukázaly, že černí koráli patří k nejpomaleji rostoucím hlubokomořským korálům. Sherwood a Edinger (2009) uvádějí odhady stáří černého korálu Stauropathes arctica až 82 kalendářních let. Prouty et al. (2011) použili radiouhlík k vytvoření odhadů stáří živých kolonií Leiopathes sp. až do 2100 let. Carreiro-Silva et al. (2013) určili stáří azorské kolonie Leiopathes sp. na 2320 let. Jedinec Leiopathes annosahas z Havaje vykazoval potenciál dosáhnout délky života přesahující 4000 let (Roark et al., 2009).
Tyto nové údaje o stáří a růstu choulostivého hlubokomořského černého korálu B. patula zdůrazňují, že se jedná o dlouhověký druh, který, jak se ukázalo v této studii, dosahuje stáří přes 385 let a vykazuje pomalé lineární a radiální tempo růstu. Výsledky naznačují pravděpodobně nízkou odolnost a pomalou dobu obnovy po narušení/odstranění. Tento výzkum bude využit pro budoucí hodnocení rizik a strategie řízení v rámci novozélandského managementu korálů (Freeman a Cryer, 2019). Pomalý růst a extrémní dlouhověkost dále poukazují na to, že tito koráli budou velmi zranitelní vůči podmořským disturbancím, jako je rybolov a těžba, a může trvat staletí, než se z antropogenních vlivů zotaví (Hitt et al., 2020).
Závěr
Stáří a rychlost růstu vzorků hlubokomořských černých korálů B. patula ze dvou oblastí Nového Zélandu, Bay of Plenty a Chatham Rise, byly stanoveny pomocí dvou různých metod; počítání růstových kroužků (sklerochronologie) a radiokarbonového datování. K počítání růstových kroužků byly použity dva různé protokoly s použitím tmavých a světlých růstových vrstev. Tyto metody poskytly rozdílné výsledky, které se rovněž neshodovaly s radiouhlíkovým stářím několika těchto korálů. Radiouhlíková stáří korálu B. patula naznačují, že tento druh korálu žije dlouho, stáří přesahuje 385 let, s pomalou lineární (5,2-9,6 mm/rok) a radiální (11,1-35,7 μm/rok) rychlostí růstu. Tyto výsledky jsou srovnatelné se stářím a rychlostí růstu jiných rodů černých korálů ze stejných oblastí Nového Zélandu i celosvětově. Stáří a rychlost růstu hlubokomořských druhů jsou rozhodující pro určení potenciálního rizika těchto organismů pro antropogenní narušení. Tyto výsledky budou použity pro budoucí hodnocení rizik a strategie řízení v rámci managementu korálů na Novém Zélandu.
Prohlášení o dostupnosti dat
Všechny soubory dat vytvořené pro tuto studii jsou součástí článku/přílohového materiálu.
Příspěvky autorů
PM, DT, HB a NH se podíleli na koncepci a návrhu studie. PM provedl veškerou přípravu vzorků, obrazovou analýzu řezů, vypracování protokolu tenkých řezů a počítání zón a mikromletí radiouhlíkových vzorků. NH provedl analýzu radiouhlíkových vzorků. SF vytvořil opravené odhady stáří ze surových radiouhlíkových dat a napsal části rukopisu. PM a DT napsali první návrh rukopisu. Všichni autoři se podíleli na revizi rukopisu.
Financování
Tento výzkum byl financován z druhého roku programu Conservation Services Programme (CSP), Department of Conservation (DOC) Project POP2017-07 Objective to „Develop a methodology to determine the age and growth characteristics of key high-risk New Zealand deep-sea (cold-water) coral species“. Finanční prostředky na úhradu publikačního poplatku pro tento časopis poskytla NIWA, Nový Zéland.
Konflikt zájmů
Autoři prohlašují, že výzkum byl prováděn bez jakýchkoli komerčních nebo finančních vztahů, které by mohly být chápány jako potenciální střet zájmů.
Poděkování
Děkujeme skupině Conservation Services Programme Group, Department of Conservation New Zealand, zejména Ianu Angusovi, Krisu Rammovi a také Freydis Hjorvarsdottir (dříve CSP, nyní Ministry of Primary Industries New Zealand) za podporu práce. Děkujeme Helen Neilové (NIWA) a Danu Sinclairovi (VUW), za podporu při poskytování jejich předběžných údajů. Děkujeme pracovníkům NIWA Sadie Millsové a Dianě Macphersonové za jejich podíl na výběru vzorků, Owenu Andersonovi za vytvoření obrázku 1, Peteru Hornovi za poskytnutí důkladné interní recenze a Rosemary Hurstové za užitečné připomínky a rady k rukopisu. Děkujeme Jeremymu Horowitzovi (James Cook University, Austrálie) za identifikaci vzorků a Dennisi Opreskovi (emeritní pracovník Smithsonian Institute) za připomínky k revizi taxonomie černých korálů. Nakonec děkujeme třem recenzentům, tento dokument měl velký přínos z jejich podrobného a konstruktivního posouzení.
Adkins, J. F., Griffin, S., Kashgarian, M., Cheng, H., Druffel, E. R. M., Boyle, E. A., et al. (2002). Radiocarbon dating of deep-sea corals. Radiocarbon 44, 567-580. doi: 10.1017/s0033822200031921
CrossRef Full Text | Google Scholar
Althaus, F., Williams, A., Schlacher, T. A., Kloser, R. J., Green, M. A., Barker, B. A., et al. (2009). Dopady lovu vlečnými sítěmi při dně na hlubinné korálové ekosystémy podmořských hor jsou dlouhodobé. Mar. Ecol. Prog. Ser. 397, 279-294. doi: 10.3354/meps08248
CrossRef Full Text | Google Scholar
Alves, E. Q., Macario, K., Ascough, P., and Bronk Ramsey, C. (2018). Celosvětový efekt mořských radiouhlíkových zásobníků: definice, mechanismy a perspektivy. Rev. Geophys. 56, 278-305. doi: 10.1002/2017RG000588
CrossRef Full Text | Google Scholar
Anderson, O., Tracey, D., Bostock, H., Williams, M., and Clark, M. (2014). Refined Habitat Suitability Modelling for Protected Coral Species in the New Zealand EEZ (Zpřesněné modelování vhodnosti stanovišť pro chráněné korálové druhy ve výlučné hospodářské zóně Nového Zélandu). Zpráva klienta NIWA vypracovaná pro Ministerstvo ochrany přírody. WLG2014-69No. 46. Auckland: NIWA.
Google Scholar
Andrews, A. H., Cailliet, G. M., Kerr, L. A., Coale, K. H., Lundstrom, C., and DeVogelaere, A. P. (2005). „Investigations of age and growth for three deep-sea corals from the Davidson Seamount off central California,“ in Cold-Water Corals and Ecosystems, eds A. Freiwald and J. M. Roberts (Berlin: Springer-Verlag), 1021-1038. doi: 10.1007/3-540-27673-4_51
CrossRef Full Text | Google Scholar
Andrews, A. H., Cordes, E. E., Mahoney, M. M., Munk, K., Coale, K. H., Cailliet, G. M. a další (2002). Age, growth and radiometric age validation of a deep-sea, habitat-forming gorgonian (Primnoa resedaeformis) from the Gulf of Alaska. Hydrobiologia 471, 101-110.
Google Scholar
Anthony, K. R. N., and Marshall, P. A. (2009). Korálové útesy a změna klimatu. Marine Climate Change Impacts and Adaptation Report Card for Australia 2009. Gland: IUCN.
Google Scholar
Bronk Ramsey, C. (2001). Vývoj radiouhlíkového kalibračního programu OxCal. Radiocarbon 43, 355-363. doi: 10.1017/s0033822200038212
CrossRef Full Text | Google Scholar
Cairns, S., Gershwin, L., Brook, F., Pugh, P., Dawson, E., Ocanþa, O., et al. (2009). „Phylum cnidaria: corals, medusae, hydroids myxozo-ans,“ in New Zealand Inventory of Biodiversity, Volume 1: Kingdom Animalia, red. D. P. Gordon (New Zealand: Canterbury University Press), 59-101.
Google Scholar
Carreiro-Silva, M., Andrews, A. H., Braga-Henriques, A., de Matos, V., Porteiro, F. M., and Santos, R. S. (2013). Variabilita v rychlosti růstu dlouhověkých černých korálů Leiopathes sp. z Azorských ostrovů. Mar. Ecol. Progr. Ser. 473, 189-199. doi: 10.3354/meps10052
CrossRef Full Text | Google Scholar
Chiswell, S. M., Bostock, H. C., Sutton, P. J. H., and Williams, M. J. M. (2015). Physical oceanography of the deep seas around New Zealand: a review [Fyzikální oceánografie hlubokých moří v okolí Nového Zélandu: přehled], New Zealand. J. Mar. Freshw. Res. 49, 1-32. doi: 10.1080/00288330.2014.992918
CrossRef Full Text | Google Scholar
Clark, M., Bowden, D., Baird, S., and Stewart, R. (2010). Effects of fishing on the benthic biodiversity of seamounts of the „Graveyard“ complex, northern Chatham Rise [Vliv rybolovu na bentickou biodiverzitu podmořských hor komplexu „Hřbitov“, severní část Chathamské výšiny]. New Zeal. Aqu. Environ. Biodivers. Rep. 46:40.
Google Scholar
Clark, M., and Rowden, A. (2009). Effects of deepwater trawling on the macro-invertebrate assemblages of seamounts on the Chatham Rise, New Zealand [Vliv hlubokomořského rybolovu vlečnými sítěmi na společenstva bezobratlých podmořských hor na Chatham Rise na Novém Zélandu]. Deep Sea Res. I 56, 1540-1554. doi: 10.1016/j.dsr.2009.04.015
CrossRef Full Text | Google Scholar
Clark, M., Tracey, D., Anderson, O., and Parker, S. (2014). Pilotní hodnocení ekologických rizik pro chráněné korály. Wellington: National Institute of Water and Atmospheric Research, 32.
Google Scholar
Clark, M. R., Bowden, D. A., Rowden, A. A., and Stewart, R. (2019). Little evidence of benthic community resilience to bottom trawling on seamounts after 15 years [Málo důkazů o odolnosti bentických společenstev vůči vlečení vlečných sítí na podmořských horách po 15 letech]. Front. Mar. Sci. 6:63. doi: 10.3389/fmars.2019.00063
CrossRef Full Text | Google Scholar
Coleman, D. C. (1991). Isotopové techniky uhlíku. Cambridge, MA: Academic Press.
Google Scholar
Consalvey, M., MacKay, K., and Tracey, D. (2006). Přehled informací o chráněných hlubokomořských korálových druzích v oblasti Nového Zélandu. Zpráva pro klienty NIWA. WLG2006-85. 60. Auckland: NIWA.
Google Scholar
Druffel, E. R. M., Griffin, S., Witter, A., Nelson, E., Southon, J., Kashgarian, M., et al. (1995). Gerardia: bristlecone Pine of the deep-sea? Geochim. Cosmochim. Acta 59, 5031-5036. doi: 10.1016/0016-7037(95)00373-8
CrossRef Full Text | Google Scholar
Druffel, E. R. M., King, L. L., Belastock, R. A., and Buesseller, K. O. (1990). Rychlost růstu hlubokomořského korálu pomocí 210Pb a dalších izotopů. Geochim. Cosmochim. Acta 54, 1493-1500.
Google Scholar
Fallon, S. J., Fifield, L. K., and Chappell, J. M. (2010). The next chapter in radiocarbon dating at the Australian National University: status report on the single stage AMS (Další kapitola radiouhlíkového datování na Australské národní univerzitě: zpráva o stavu jednostupňového AMS). Nuclear Instr. Methods Phys. Res. Sec. B 268, 898-901. doi: 10.1016/j.nimb.2009.10.059
CrossRef Full Text | Google Scholar
Freeman, D., and Cryer, M. (2019). „Current management measures and threats,“ in The State of Knowledge of Deep-Sea Corals in the New Zealand Region, Chap. 9, eds D. M. Tracey and F. Hjorvarsdottir (Auckland: NIWA Science and Technology Series).
Google Scholar
Gammon, M. J., Tracey, D. M., Marriott, P. M., Cummings, V. J., and Davy, S. K. (2018). The physiological response of the deep-sea coral Solenosmilia variabilis to ocean acidification (Fyziologická reakce hlubokomořského korálu Solenosmilia variabilis na okyselování oceánu). PEERJ 6:e5236. doi: 10.7717/peerj.5236
PubMed Abstract | CrossRef Full Text | Google Scholar
Griffin, S., and Druffel, E. M. (1989). Zdroje uhlíku pro hlubokomořské korály. Radiocarbon 31, 533-543. doi: 10.1017/s0033822200012121
CrossRef Full Text | Google Scholar
Hitt, N. T., Sinclair, D. J., Fallon, S. J., Neil, H. L., Tracey, D. M., Komugabe-Dixson, A., et al. (2020). Growth and longevity of New Zealand black corals [Růst a dlouhověkost novozélandských černých korálů]. Deep Sea Res. I 2020:103298. doi: 10.1016/j.dsr.2020.103298
CrossRef Full Text | Google Scholar
Komugabe, A. F., Fallon, S. J., Thresher, R. E., and Eggins, S. M. (2014). Moderní stáří povrchových nádrží Tasmánského moře z hlubokomořských černých korálů. Deep-Sea Res. 99, 207-212. doi: 10.1016/j.dsr2.2013.05.024
CrossRef Full Text | Google Scholar
Komugabe-Dixson, A. F., Fallon, S. J., Eggins, S. M., and Thresher, R. E. (2016). Radiocarbon evidence for mid-late Holocene changes in southwest Pacific Ocean circulation (Radiouhlíkové důkazy změn cirkulace v jihozápadním Pacifiku v polovině pozdního holocénu). Paleoceanography 31, 971-985. doi: 10.1002/2016PA002929
CrossRef Full Text | Google Scholar
Love, M. S., Yoklavich, M. M., Black, B. A., and Andrews, A. H. (2007). Age of black coral (Antipathes dendrochristos) colonies, with notes on associated invertebrate species [Stáří kolonií korálů černých (Antipathes dendrochristos) s poznámkami k přidruženým druhům bezobratlých]. Bull. Mar. Sci. 80, 391-400.
Google Scholar
Matsumoto, K. (2007). Radiouhlíkové stáří cirkulace světových oceánů. J. Geophys. Res. 112:C09004. doi: 10.1029/2007JC004095
CrossRef Full Text | Google Scholar
Mortensen, P. B., and Buhl-Mortensen, L. (2005). Morfologie a růst hlubokovodních gorgonií Primnoa resedaeformis a Paragorgia arborea. Mar. Biol. 147, 775-788. doi: 10.1007/s00227-005-1604-y
CrossRef Full Text | Google Scholar
Noé, S. U., Lembke-Jene, L., and Chr Dullo, W. (2008). Varying growth rates in bamboo corals: sclerochronology and radiocarbon dating of a mid-Holocene deep-water gorgonian skeleton (Keratosis sp.: Octocorallia) z Chatham Rise (Nový Zéland). Facies 54, 151-166. doi: 10.1007/s10347-007-0129-x
CrossRef Full Text | Google Scholar
Pinkerton, M. H. (2016). Satelitní pozorování barvy oceánu a fytoplanktonu ve výlučné ekonomické zóně Nového Zélandu v letech 1997-2016. Ministerstvo životního prostředí. Wellington: NIWA.
Google Scholar
Prouty, N. G., Fisher, C. R., Demopoulous, A. W. J., and Druffel, E. R. M. (2016). Growth rates and age of dee-sea corals impacted by the Deepwater Horizon oil spill (Rychlost růstu a stáří dee-mořských korálů zasažených ropnou skvrnou Deepwater Horizon). Deep-Sea Res. II 129, 196-212. doi: 10.1016/j.dsr2.2014.10.021
CrossRef Full Text | Google Scholar
Prouty, N. G., Roark, E. B., Buster, N. A., and Ross, S. W. (2011). Growth rate and age distribution of deep-sea black corals in the Gulf of Mexico. Mar. Ecol. Prog. Ser. 423, 101-115. doi: 10.3354/meps08953
CrossRef Full Text | Google Scholar
Reimer, P. J., Bard, E., Bayliss, A., Beck, J. W., Blackwell, P. G., Bronk Ramsey, C., et al. (2013). Kalibrační křivky radiouhlíkového stáří IntCal13 a Marine13 0-50 000 let cal BP. Radiocarbon 55, 1869-1887. doi: 10.2458/azu_js_rc.55.16947
PubMed Abstract | CrossRef Full Text | Google Scholar
Reimer, P. J., and Reimer, R. W. (2001). Databáze korekcí mořských nádrží a on-line rozhraní. Radiocarbon 43, 461-463. doi: 10.1017/s0033822200038339
CrossRef Full Text | Google Scholar
Roark, E. B., Guilderson, T. P., Dunbar, R. B., Fallon, S. J., and Mucclarone, D. A. (2009). Extrémní dlouhověkost u bílkovinných hlubokomořských korálů. Proc. Natl. Acad. Sci. U.S.A. 106, 5204-5208. doi: 10.1073/pnas.0810875106
PubMed Abstract | CrossRef Full Text | Google Scholar
Roark, E. B., Guilderson, T. P., Dunbar, R. B., and Ingram, B. L. (2006). Radiocarbon based ages and growth rates [Stáří na základě radiouhlíku a rychlost růstu]: Hawaiian deepsea corals. Mar. Ecol. Progr. Ser. 327, 1-37.
Google Scholar
Sánchez, J., and Rowden, A. (2006). „Octocoral diversity on New Zealand Seamounts,“ in Proceedings of the Presented at the 10th International Coral Reef Symposium, Okinawa.
Google Scholar
Sherwood, O. A., and Edinger, E. N. (2009). Ages and growth rates of some deep-sea gorgonian and antipatharian corals of Newfoundland and Labrador [Stáří a rychlost růstu některých hlubokomořských gorgoniových a antipatrových korálů Newfoundlandu a Labradoru]. Can. J. Fish. Aquatic Sci. 66, 142-152. doi: 10.1139/f08-195
CrossRef Full Text | Google Scholar
Sherwood, O. A., Scott, D. B., Risk, M. J., and Guilderson, T. P. (2005). Radiocarbon evidence for annual growth rings in the deep-sea octocoral Primnoa resedaeformis (Radiouhlíkové důkazy ročních růstových kruhů u hlubokomořského chobotnatce Primnoa resedaeformis). Mar. Ecol. Prog. Ser. 301, 129-134. doi: 10.3354/meps301129
CrossRef Full Text | Google Scholar
Sikes, E., Burgess, S., Grandpre, R., and Guilderson, T. (2008). Assessing modern deep-water age in the New Zealand region using deep-water corals [Posuzování moderního stáří hlubokých vod v oblasti Nového Zélandu pomocí hlubokomořských korálů]. Deep Sea Res. I 55, 38-49. doi: 10.1016/j.dsr.2007.10.004
CrossRef Full Text | Google Scholar
Stuiver, M., and Polach, H. A. (1977). Diskusní hlášení údajů o 14C. Radiocarbon 19, 355-363. doi: 10.1017/S0033822200003672
CrossRef Full Text | Google Scholar
Thresher, R., Rintoul, S. R., Koslow, J. A., Weidman, C., Adkins, J., and Proctor, C. (2004). Oceanic evidence of climate change in southern Australia over the last three centuries (Oceánské důkazy klimatických změn v jižní Austrálii za poslední tři století). Geophys. Res. Lett. 31:l07212.
Google Scholar
Thresher, R. E., MacRae, C. M., Wilson, N. C., and Gurney, R. (2007). Vliv prostředí na složení kostry hlubokovodních gorgonií (Keratoisis spp.; Isididae). Bull. Mar. Sci. 8, 409-422.
Google Scholar
Tracey, D., Bostock, H., and Shaffer, M. (2018). Metody stárnutí chráněných hlubokomořských korálů: A Review and Recommendation for an Ageing Study (Přehled a doporučení pro studii stárnutí). DOC Contract 4527 GMC – Age and Growth of coral (POP2017-07). Zpráva klienta NIWA WN2018035. Auckland: NIWA, 40.
Google Scholar
Tracey, D., Neil, H., Gordon, D., a O’Shea, S. (2003). Chronicles of the deep: ageing deep-sea corals in New Zealand waters (Kroniky hlubin: stárnutí hlubokomořských korálů ve vodách Nového Zélandu). NIWA Water Atmosph. 11, 22-24.
Google Scholar
Tracey, D. M., a Hjorvarsdottir, F. (eds) (2019). „The state of knowledge of deep-sea corals in the new zealand region,“ in NIWA Science and Technology Series Number 84, (Auckland: NIWA),140.
Google Scholar
Tracey, D. M., Neil, H., Marriott, P., Andrews, A. H., Cailliet, G. M., and Sanchez, J. A. (2007). Age and growth of two genera of deep-sea bamboo corals (family Isididae) in New Zealand waters. Bull. Mar. Sci. 81, 393-408.
Google Scholar
Williams, A., Schlacher, T. A., Rowden, A. A., Althaus, F., Clark, M. R., Bowden, D. A., et al. (2010). Seamount megabenthic assemblages fail to recover from trawling impacts (Podmořská megabentická společenstva se nedokáží zotavit z dopadů vlečných sítí). Mar. Ecol. 31, 183-199. doi: 10.1111/j.1439-0485.2010.00385.x
CrossRef Full Text | Google Scholar
Williams, B., and Grottoli, A. G. (2010). Proměnlivost stabilních izotopů dusíku a uhlíku (δ15N a δ13C) u měkkých korálů a taxonů černých korálů v mělkém tropickém Pacifiku a důsledky pro paleoceánografické rekonstrukce. Geochim. Cosmochim. Acta 74, 5280-5288. doi: 10.1016/j.gca.2010.06.026
CrossRef Full Text | Google Scholar
Williams, B., Risk, M. J., Ross, S. W., and Sulak, K. J. (2007). Údaje o stabilních izotopech z hlubokovodních antipatogenů: Záznamy za 400 let z jihovýchodního pobřeží Spojených států amerických. Bull. Mar. Sci. 81, 437-447.
Google Scholar
.
Leave a Reply