Frontiers in Plant Science
De olika typerna av heterokroni
På 1870-talet identifierade Ernst Haeckel tidsmässiga och rumsliga förändringar i utvecklingen hos en ättling i förhållande till dess förfader som de två viktigaste mekanismerna för evolutionen (Haeckel, 1875). Haeckel kallade spatiala förändringar för heterotopi och temporala förändringar för heterokroni.
Däremot har betydelsen av heterokroni förändrats sedan Haeckel först myntade begreppet. Haeckel använde termen heterokroni för att hänvisa till avvikelser från hans välkända ”Biogenetiska lag”, som säger att sekvensen av utvecklingshändelser (ontogeni) i stort sett återger sekvensen av händelser i artens evolutionära historia (fylogeni) (Haeckel, 1875). Heterokroni hänvisade alltså ursprungligen till en förändring i tidpunkten för när ett särdrag uppträder i en organisms utvecklingssekvens i förhållande till den sekvens som förekom i organismens fylogeni. I mitten av 1900-talet kopplade De Beer (1951) bort heterokroni från rekapitulation. Han använde heterokroni för att beteckna skillnader i tidpunkten för utvecklingshändelser när han jämförde två besläktade arter, för att förklara hur heterokroni kan generera mångfald bland organismer. Gould (1977) kopplade återigen begreppet heterokroni till rekapitulation och definierade heterokroni som ”förändringar i den relativa tidpunkten för uppkomsten och utvecklingshastigheten för karaktärer som redan finns hos förfäderna”, och betonade förändringar i den relativa storleken och formen, snarare än i tidpunkten för utvecklingshändelser, för att upptäcka heterokroni. Vid sekelskiftet 2000 föreslog Smith att det skulle vara mer användbart att definiera två typer av heterokroni: ”tillväxtheterokroni”, som betonar förändringar i slutlig storlek och form, och ”sekvensheterokroni”, som ligger närmare Haeckels och de Beers ursprungliga användning, och som gör det möjligt att förklara fenotypisk variation genom förändringar i tidpunkten för utvecklingshändelser (Smith, 2002, 2003; Keyte och Smith, 2014).
Förloppsheterokroni (nedan kallad ”heterokroni”) kan delas in i två kategorier: paedomorfos och peramorfos. Jämfört med förfädernas utveckling resulterar paedomorfos i ett juvenilt eller enkelt resultat, medan peramorfos resulterar i en vuxen eller mer komplex fenotyp. Var och en av dessa två kategorier av heterokroni kan vara resultatet av variation i tidpunkten för start, förskjutning eller hastighet av en utvecklingsprocess, enligt Alberch et al. (1979). Denna variation kan resultera i sex olika typer av heterokroni (figur 1). Paedomorfos kan bero på att en utvecklingsprocess avslutas för tidigt (progenesis), att processen startar för sent (postdisposition) eller att utvecklingstakten är långsammare (neoteny). Peramorfos är resultatet av en förlängd utvecklingsperiod på grund av ett senare slut (hypermorfos) eller en tidigare start (pre-displacement), eller av en högre utvecklingstakt (acceleration) (figur 1A). Hypotetiska exempel på peramorfos och paedomorfos i växternas embryogenes och vegetativa utveckling visas i figurerna 1B,C.
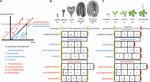
FIGUR 1. Typer av heterokroni. (A) Schematisk framställning av de 6 typerna av heterokroni. Den svarta linjen representerar den tid som krävs för att nå ett visst utvecklingsstadium i den förfädernas ontogeni. Blå linjer visar de 3 typerna av paedomorfos: progenesis (tidig förskjutning), postdisplacement (fördröjd förskjutning) och neoteny (långsammare utvecklingstakt). Röda linjer visar de tre typerna av peramorfos: hypermorfos (fördröjd förskjutning), pre-displacement (tidig start) och acceleration (högre utvecklingstakt). Teckningen i (A) är baserad på Alberch et al. (1979) och Geuten och Coenen (2013). (B) Scenarier för heterokroni för embryogenes. Arabidopsis embryogenes tas här som den hypotetiska förfädersutvecklingen, uppdelad i fyra stadier i illustrativt syfte. Ett femte stadium, där embryot har producerat de två första bladen plus de två kotyledonen (betecknas embryonal fröplanta), föreslås som det sista stadiet i den peramorfa embryogenesen, medan den paedomorfa embryogenesen förväntas avslutas vid hjärtstadiet. (C) Heterokroniska scenarier för vegetativ utveckling. I detta fall representerar Arabidopsis vegetativa utveckling, där växten fortsätter att producera cauline/reproduktiva blad på stammen före blomningen (ritad överst), ett hypotetiskt fall av peramorf vegetativ utveckling, jämfört med en hypotetisk förfader som blommar i vuxenstadiet utan att producera caulineblad. Paedomorf vegetativ utveckling förutspås resultera i växter som blommar i det unga stadiet.
Men även om dessa klassificeringar är användbara för att illustrera förändringar i utvecklingstidpunkten är det ofta svårt att skilja mellan olika typer av heterokroni när man tittar på exempel från det verkliga livet. I många fall kan evolutionen resultera i olika typer av heterokroni, som var och en uppträder i ett diskret skede av en utvecklingsprocess (granskad i växter av Li och Johnston, 2000).
Heterokroni i växternas evolution och diversifiering
Landväxter (embryofyter) har genomgått många morfologiska innovationer sedan de uppkom i mitten av den ordoviciska perioden, för cirka 470 miljoner år sedan. Tidiga divergerande släktskap uppstod under en period på mer än 100 miljoner år, under silur och tidig devon (Kenrick och Crane, 1997; Pires och Dolan, 2012; Harrison och Morris, 2018). Embryofyter utvecklades från en sötvattensalgsförfader som var besläktad med de existerande charofytgrupperna Charales, Coleochaetales och Zygnematales. Övergången till tillväxt på land, som observerades hos bryofyter (hornvårtor, mossor och levermossor), innebar ursprunget av sporer, växling av generationer av gametofyter och sporofyter, enaxliga former och tredimensionell tillväxt. Innovationer hos terrestra kärlväxter (lycofyter, monilofyter och spermatofyter) omfattade bifurkation, obestämdhet, sporofytisk dominans, axillär förgrening och bildandet av meristem, blad och rötter (Harrison, 2017).
Flera av de evolutionära stegen ovan har kopplats till heterokroni. När det gäller sporogenesens ursprung tyder data från bryofyter på att sporer producerades direkt från zygoter i en process som innefattar tidig cytokinesis, acceleration av meiosen och fördröjd väggdeposition från zygoten till meiosporerna (Brown och Lemmon, 2011). Den grenade sporofyten (polisporangiofyten) har antagits ha utvecklats genom förlängd vegetativ tillväxt av den apikala cellen. Denna längre period av vegetativ tillväxt föreslogs resultera i en förlängd embryonal axel, skottförgrening och en fördröjning av övergången till reproduktiv tillväxt, som producerar sporangiet (Rothwell et al., 2014; Tomescu et al., 2014). Ytterligare studier av växtfossil bör ge fler bevis för forntida växtmorfologier, vilket skulle möjliggöra jämförelser mellan samtida och utdöda former (Rothwell et al., 2014).
I utdöda växter har heterokroniska förändringar identifierats i gametofytutvecklingen, embryogenesen, den vegetativa utvecklingen, skottmognaden och den florala morfogenesen. Den kvinnliga gametofyten hos Gnetum skiljer sig strukturellt från andra växter på grund av skillnader i tidpunkten för befruktning och somatisk utveckling. Temporala förändringar i cellcykelutvecklingen har bidragit till diversifierade tidsmönster för spermatogenes och gametfusion under befruktning (Friedman, 1999; Tian et al., 2005). I ett fall av progenesering sker befruktningen i ett fritt kärnstadium av den somatiska utvecklingen, ett juvenilt stadium jämfört med den förfädernas somatiska ontogeni, vilket utesluter differentiering av äggceller (Friedman och Carmichael, 1998). Den apomiktiska utvecklingen av Boechera äggceller har förknippats med heterokroniska genuttrycksmönster jämfört med icke-apomiktiska (sexuella) äggceller (Sharbel et al., 2010). Utvecklingen av Rafflesiaceae, en holoparasitisk växtfamilj som infekterar vinstockar, uppvisar två heterokroniska förändringar: ett stopp i proembryonstadiet, som kan betraktas som ett exempel på neoteni, och en acceleration av övergången från den odifferentierade endofyten till blomning, där man hoppar över den vegetativa skottmognaden (Nikolov et al, 2014).
En QTL-analys (Quantitative Trait Loci) som jämför Eucalyptus globulus-populationer med tidig förändring av den vegetativa fasen och populationer där förändringen av den vegetativa fasen är fördröjd flera år identifierade uttrycket av mikroRNA:t EglMIR156.5 som ansvarigt för den heterokroniska variationen av förändringen av den vegetativa fasen hos E. globulus (Hudson et al., 2014). I en annan QTL-analys drogs slutsatsen att heterokroni ligger till grund för den naturliga variationen i Cardamine hirsuta bladform (Cartolano et al., 2015). QTL-mappning fastställde att effekten orsakas av cis-regulatorisk variation i den florala repressorn ChFLC så att populationer med lågexpressiva ChFLC-alleler uppvisar både tidig blomning och accelererande förvärv av vuxna bladegenskaper, särskilt ökat antal bladblad. Morfometriska och QTL-analyser har fastställt att heterokroniska mutationer bidrar till den naturliga variationen hos Antirrhinum och till druvornas heteroblast (Costa et al., 2012; Chitwood et al., 2016). En Principal Component Analysis (PCA) av de ontogenetiska banorna för bladformen bland de tre släktena av marsialiska ormbunkar (Marsilea, Regnellidium och Pilularia) föreslog att de uppvisar en paedomorf fenotyp, jämfört med den mer komplexa förfädersutvecklingen, som orsakas av accelererad tillväxthastighet och tidig avslutning vid en förenklad bladform (Pryer och Hearn, 2009).
Evolutionär mångfald i blomställningens arkitektur hos Solanaceae moduleras av heterokroniska skiftningar i förvärvet av blommans öde (Lippman et al, 2008; Park et al., 2012). En jämförelse av transkriptom av meristemmognad från fem domesticerade och vilda Solanaceae-arter avslöjade en topp av uttrycksdivergens, som liknar ”omvänt timglas”-modellen för djurembryogenes, enligt vilken en divergensperiod i mitten av utvecklingen driver den morfologiska variationen (Lemmon et al., 2016). Hos gräs resulterar en fördröjning av övergången från skottmeristem (SM) till blommeristem (FM) i mer komplexa pannor (Kyozuka et al., 2014). Popplar (Populus sp.) och pilar (Salix sp.) bär kompakta enkönade blomställningar som kallas ”catkins”, vilka har föreslagits vara utvecklade från en förenkling av pannlikationsformen genom en tidig SM- till FM-övergång (Cronk et al., 2015). Heterokroniska förändringar har också bidragit till naturlig variation i blomningstid och skottarkitektur bland Mimulus guttatus-populationer (Baker och Diggle, 2011).
Morfologisk mångfald av perianth i Dipsacoideae orsakas av heterokroniska förändringar i organinitiering, specifikt i antalet sepaler (Naghiloo och Claßen-Bockhoff, 2017). Den stora formdiversiteten hos sepalerna bland Iris-arter beror mer på heterokroni än på heterotopiska förändringar (Guo, 2015). En studie i Brassicaceae visade att evolutionen av korollamonosymmetri från den polysymmetriska förfädersblomman innebar en heterokronisk förskjutning av uttrycket av CYC2-gener (en klad av TCP-transkriptionsfaktorer) från ett tidigt adaxialt uttryck i förfädersblommans meristem till ett senare adaxialt uttryck vid kronbladets utveckling (Busch et al., 2012). Heterokroniskt, men inte heterotopiskt, CYC2-uttryck har också förknippats med en förlust av papillerade koniska celler i kronbladen och en övergång till ett system med fågelpollinering hos Lotus (Ojeda et al., 2017). En paedomorfisk morfologi, där blommorna håller mogna pollen i oöppnade knoppliknande strukturer, ledde till specialiserad pollinering i en klad av Madagaskars vinrankor (Euphorbiaceae) (Armbruster et al., 2013). Utvecklingen av cleistogama capitulum från ett chasmogamt förfäders tillstånd är ett klassiskt exempel på pedomorfos, eftersom den cleistogama formen uppvisar juvenila drag (Lord och Hill, 1987). Cleistogami hos Asteraceae utvecklades specifikt genom förskjutning och progenesering av blomutvecklingen, samt neoteni av alla andra virvlar än gynoecium (Porras och Muñoz, 2000). Mångfalden av blommors morfologier inom Jaltomata, ett Solanaceae-släkte, beror på hypermorfosering och acceleration av vissa korolla-egenskaper (Kostyun et al., 2017). Mångfalden hos Azorernas fjärilsorkidéer orsakas också av florala heterokroniska skiftningar (Bateman et al., 2014). Nyligen betonade Ronse de Craene (2018) betydelsen av heterokroni i tre utvecklingsprocesser: phyllotaxis, utvecklingen av gemensamma primordier av ståndare och blad och obdiplostemoni, genom att koppla samman förändringar i tillväxthastighet med fördröjd organinitiering. Heterokroniska tillväxthastigheter hos perianth och stil, och tidig hypanthiumförlängning, är ansvariga för den stora artdiversiteten inom det morfologiskt homogena Eugenia-släktet (Vasconcelos et al., 2018). Slutligen är heterokroniskt uttryck av fw2.2-allelen, som påverkar celldelningen i den tidiga fruktutvecklingen, ansvarig för den naturliga variationen i tomatfruktstorlek (Cong et al., 2002).
Studier av heterokroniska mutanter för att belysa genetisk kontroll av timing
Studier av mutanter som påverkas i utvecklingstidpunkten har kastat ljus över genetiska vägar som kontrollerar morfogenes och utvecklingsövergångar. Heterokroni kan orsakas av tidigare eller senare aktivering eller repression av dessa vägar.
Mutanter för leafy cotyledon (lec), dicer-like1 (dcl1) och extra cotyledon (xtc) hos Arabidopsis thaliana representerar heterokroniska fenotyper som har hjälpt till att definiera program för fröets mognad. lec-mutanter ger upphov till cotyledon med egenskaper som liknar bladidentitet (Meinke, 1992), vilket är ett tydligt exempel på homeos: att en struktur ersätts av en annan. Det är dock ofta svårt att skilja mellan homeos och heterokroni, eftersom homeos kan vara resultatet av både heterokroni och heterotopi (Li och Johnston, 2000; Geuten och Coenen, 2013). Under sen embryogenes främjar LEC2 frömognad och undertrycker postembryonal identitet (Stone et al., 2008). Liksom lec-mutanter uppvisar dcl1-mutanter peramorfiska fenotyper under embryogenesen, eftersom kloroplastutvecklingen och genuttrycket för frölagringsprotein inträffar tidigare än i vildtypembryon. DCL1 krävs för biogenes av mikroRNA, som undertrycker frömognad genom huvudregulatorerna LEC2 och FUSCA3 (Nodine och Bartel, 2010; Willmann et al., 2011). Mutanterna xtc1, xtc2 och altered meristem programming1 (amp1) uppvisar en homeotisk fenotyp där de första ett eller två bladen förvandlas till kotyledoner (Conway och Poethig, 1997). Hos dessa tre mutanter fördröjs den globulära till hjärtövergången, vilket orsakar ett förstorat skottmeristem, vilket leder till extra organbildning under embryogenesen. Denna fenotyp kan tolkas som hypermorfos, en typ av peramorfos, eftersom embryoutvecklingen förlängs till en mer utvecklad form (som det peramorfa scenariot i figur 1B). Men om vegetativ utveckling väljs som referenspunkt skulle lec-mutanter representera ett fall av förflyttning (peramorfos), och xtc1, xtc2 och amp1 skulle representera efterflyttning (paedomorfos) i förvärvet av bladidentitet. AMP1 krävs för miRNA-medierad translationsrepression på ER-membranet (Li et al., 2013).
Genetisk reglering av övergången från juvenil till vuxen, även kallad vegetativ fasförändring, har studerats med hjälp av både paedomorfiska och peramorfiska mutanter. Screening av mutanter som visar en tidig vuxen (peramorf) fenotyp producerade alleler av gener relaterade till små RNA-biogenes som zippy/ago7, sgs3 och rdr6 (Hunter et al., 2003; Peragine et al., 2004), medan mutanter med en sen vuxen (paedomorf) fenotyp beror på ökade miRNA-nivåer (Gillmor et al., 2014; Xu M. et al., 2016, 2018; Guo et al., 2017). Den genetiska grunden för dessa fenotyper förklaras av förändrat uttryck av de närbesläktade mikroRNA:erna miR156 och miR157, de viktigaste regulatorerna av vegetativ fasförändring som verkar genom att undertrycka SPL:s transkriptionsfaktorer på både transkriptions- och translationsnivå, på ett tröskelberoende sätt (Wu och Poethig, 2006; Chuck et al., 2007; Poethig, 2013; He et al., 2018). Både MIR156- och SPL-genfamiljerna regleras direkt av epigenetiska markeringar: MIR156 aktiveras av H3K4me3 och H3K4ac, som främjas av SWR1-C-komplexet och kromatinremodelleraren BRAHMA, och undertrycks av H3K27me3, som främjas av Polycomb-proteinerna SWINGER och CURLY LEAF och av kromatinremodelleraren PICKLE (Xu M. et al., 2016; Xu Y. et al., 2016; Xu M. et al., 2018). SPL-gener aktiveras av histonacetylering som medieras av det SAGA-liknande komplexet och undertrycks av H2AUb som medieras av Polycomb-proteinerna RING1A och RING1B (Kim et al., 2015; Li et al., 2017). Studier av växter med förlust- eller gain-of-funktion av SPL-gener har också definierat en endogen blomningsväg där vissa SPL-gener främjar uttrycket av miR172, som i sin tur främjar blomning genom att reprimera blomningsrepressorerna i APETALA2-familjen (Wang et al., 2009; Wu et al., 2009; Huijser och Schmid, 2011).
Bortsett från större utvecklingsövergångar kan organmorfogenesen också påverkas av heterokronisk aktivering av genetiska regulatorer. Till exempel resulterar tidig groning av Brassica rapa-embryon i organ med mosaik av kotyledon- och bladidentitet (Fernandez, 1997), och differentiellt tidsmässigt uttryck av klass II av TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATION CELL FACTOR (TCP)-generna resulterar i blad med olika storlek och form (Efroni et al, 2008).
Mekanismer som driver förändringar i utvecklingstidpunkten i växter (transkriptionell, metabolisk och cellulär heterokroni)
Transkriptionell heterokroni avser en förändring i tidpunkten för aktivering eller repression av genuttryck, och orsakas ofta av förändringar i cis-regulatoriska genregioner (Pham et al., 2017). Transkriptionell heterokroni i de genetiska vägar som nämns ovan är ett vanligt sätt att producera heterokroniska fenotyper. Metabolisk kontroll av banor som reglerar utvecklingsövergångar (här kallad ”metabolisk heterokroni”) och tidsmässig kontroll av cellproliferation, celexpansion och celldifferentiering (här kallad ”cellulär heterokroni”) är andra mekanismer som driver heterokroni i växter (figur 2). Transkriptionella, metaboliska och cellulära processer är sammankopplade, så det molekylära ursprunget till heterokroni kan bero på en kombination av mekanismer.

FIGUR 2. Mekanismer som driver heterokroni hos växter. (A) Cellulär heterokroni. Sekvensen av rosettblad som produceras under den vegetativa utvecklingen visas. Fenotypen hos Arabidopsis Columbia vildtypsplantor tas som ett hypotetiskt förfäderstillstånd; ett paedomorfiskt scenario (neoteni) och ett peramorfiskt scenario (acceleration) visas också. Pilarna representerar riktningen på vågen av celldelningsstoppet i en basipetal gradient, där cellerna under den streckade linjen fortfarande delar sig aktivt medan cellerna ovanför expanderar och/eller differentieras. (B) Transkriptionell och metabolisk heterokroni. En grafisk representation av den förutspådda abundansen av mikroRNA miR156/7 och dess mål, SPL-generna, samt sockerabundansen längs tiden för den vegetativa utvecklingen, som motsvarar fenotyperna i (A). Streckade linjer representerar tröskelvärdet för miR156/7 och SPL-överflödet som leder till juvenilitet (över tröskelvärdet) eller vuxen ålder (under tröskelvärdet). I detta fall resulterar en fördröjning av repressionen av miR156 och aktivering av SPL-uttryck och sockerproduktion i paedomorfos, medan en tidig nedbrytning av miR156 och tidig aktivering av SPL-uttryck och sockerproduktion resulterar i peramorfos. Dessa teckningar är förenklade framställningar och återspeglar inte de exakta mängderna i naturen. Det faktiska mönstret för miR156/SPL-överflödet är närmare det som avbildas för tillståndet ”peramorfos”.
Metabolisk heterokroni kan påverkas av hormoner, socker och redoxsignaler (Jia et al., 2017). Differentiell biosyntes, transport och perception av hormoner som auxin, jasmonsyra (JA), gibberellin och abscisinsyra påverkar heterokroni genom att kontrollera regulatorer av utvecklingsprocesser. Mutanter i AUXIN RESPONSE FACTORS (ARF) ARF3 och ARF4 fördröjer till exempel den vuxna övergången (Fahlgren et al., 2006; Hunter et al., 2006), och auxinhomeostasen kontrollerar övergången från underhåll av blommande stamceller till gynoeciumbildning (Yamaguchi et al., 2017). Exogen JA kan fördröja den vuxna övergången genom att skjuta upp minskningen av miR156-uttrycket (Beydler et al., 2016), och gibberellin påskyndar blomningen genom att frigöra SPL-gener från repression av DELLA-proteiner (Yu et al., 2012). Näringsstatus har förknippats med kontrollen av vegetativa fasförändringar sedan början av 1900-talet (Goebel, 1908). Socker som produceras av fotosyntesen är nödvändigt för att förvärva vuxna egenskaper och är delvis ansvarig för minskningen av miR156-uttrycket i sen vegetativ utveckling (Yang et al., 2013; Yu et al., 2013; Buendía-Monreal och Gillmor, 2017). HEXOKINASE1 (HXK1) och Trehalose-6-fosfat (T6P) är viktiga för den sockermedierade repressionen av miR156, vilket främjar vegetativ fasförändring och blomning (Wahl et al., 2013; Yang et al., 2013).
På cellulär nivå består organogenesen av en sekvens av tre stadier: etableringen av polaritet, cellproliferation och celexpansion (Walcher-Chevillet och Kramer, 2016). Tidpunkten för initiering och avslutande av dessa stadier är avgörande för organens storlek och form, och heterokroni i denna sekvens resulterar i diversifiering av organens storlek och form (figur 2). Övergången från cellproliferation till cellutvidgning och differentiering kräver samordning mellan cellcykeln och celltillväxten (Sablowski och Carnier Dornelas, 2014). I blad av modellväxter rör sig denna övergång som en basipetal våg av cellcykelstopp som börjar i den distala delen av primordiet och rör sig mot basen. Cellerna bakom mitotiska arresteringsfronten blir starkt vakuoliserade och börjar expandera (Donnelly et al., 1999; Czesnick och Lenhard, 2015). Andra växtarter kan dock visa diffus tillväxt och acropetala eller dubbelriktade gradienter för cellcykelstopp (Das Gupta och Nath, 2015). Förvärvet av fotosyntetisk kapacitet krävs för övergången från celldelning till cellutvidgning (Andriankaja et al., 2012). Detta skifte korrelerar med sockrets roll när det gäller att främja Target of Rapamycin (TOR)-vägen och undertrycka Sucrose-non-fermenting1-relaterat kinas 1 (SnRK1): TOR och T6P inducerar celexpansion genom att främja makromolekylär syntes, medan SnRK1 främjar katabolism (Tsai och Gazzarrini, 2014; Sablowski, 2016). Två mikroRNA spelar motsatta roller i detta cellulära skifte: miR319 undertrycker uttrycket av klass II TCP-faktorer, som är hämmare av cellproliferation, medan miR396 begränsar uttrycket av GROWTH REGULATING FACTORS (GRFs), som fördröjer differentiering (Das Gupta och Nath, 2015; Maugarny-Calés och Laufs, 2018). Övergången från ett obestämt skott apikalt meristem till ett bestämt blommande meristem involverar också tidsmässig reglering av den cellulära identiteten. Tidpunkten för AGAMOUS aktivering av KNUCKLES, som i sin tur reprimerar WUSCHEL, definierar det tidsmässiga fönstret för obestämdhet och följaktligen storleken och antalet organ (Sun et al., 2014).
Slutsats
En förståelse för den tidsmässiga regleringen av växternas utveckling är nödvändig för att bättre kunna uppskatta mångfalden av växtformer som vi ser i naturen, för att förklara växternas morfologiska evolution och för att kunna manipulera växternas arkitektur till förmån för jordbruket. Såsom beskrivs ovan driver samverkande transkriptionella, metaboliska och cellulära mekanismer heterokroni hos existerande arter. Ytterligare forskning om dessa vägar i angiospermer och basala växtlinjer bör avslöja mer om de förändringar i utvecklingstidpunkten som har drivit utvecklingen av växternas utveckling.
Författarbidrag
MB-M utformade och skrev manuskriptet. CSG redigerade och reviderade manuskriptet.
Intressekonfliktförklaring
Författarna förklarar att forskningen utfördes i avsaknad av kommersiella eller ekonomiska relationer som skulle kunna tolkas som en potentiell intressekonflikt.
Alberch, P., Gould, S., Oster, G., and Wake, D. (1979). Size and shape in ontogeny and phylogeny. Paleobiology 5, 296-317. doi: 10.1017/S0094837300006588
CrossRef Full Text | Google Scholar
Andriankaja, M., Dhondt, S., DeBodt, S., Vanhaeren, H., Coppens, F., DeMilde, L., et al. (2012). Utgång från proliferation under bladutveckling i Arabidopsis thaliana: en inte så gradvis process. Dev. Cell 22, 64-78. doi: 10.1016/j.devcel.2011.11.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Armbruster, W. S., Lee, J., Edwards, M. E. och Baldwin, B. G. (2013). Floral paedomorfi leder till sekundär specialisering vid pollinering av Madagaskar Dalechampia (Euphorbiaceae). Evolution 67, 1196-1203. doi: 10.1111/evo.12002
PubMed Abstract | CrossRef Full Text | Google Scholar
Baker, R. L., and Diggle, P. K. (2011). Knutspecifik förgrening och heterokroniska förändringar ligger bakom skillnader på populationsnivå i Mimulus guttatus (Phrymaceae) skottarkitektur. Am. J. Bot. 98, 1924-1934. doi: 10.3732/ajb.1100098
PubMed Abstract | CrossRef Full Text | Google Scholar
Bateman, R. M., Rudall, P. J., Bidartondo, M. I., Cozzolino, S., Tranchida-Lombardo, V., Carine, M. A., et al. (2014). Artbestämning via blommig heterokroni och förmodat byte av mykorrhizal värd hos endemiska fjärilsorkidéer på Azorernas skärgård. Am. J. Bot. 101, 979-1001. doi: 10.3732/ajb.1300430
PubMed Abstract | CrossRef Full Text | Google Scholar
Beydler, B. D., Osadchuk, K., Cheng, C.-L., Manak, J. R. och Irish, E. E. (2016). I majsens juvenila fas ses uppreglering av stressresponsgener och förlängs av exogent JA. Plant Physiol. 171, 2648-2658. doi: 10.1104/pp.15.01707
PubMed Abstract | CrossRef Full Text | Google Scholar
Brown, R. C., and Lemmon, B. E. (2011). Sporer före sporofyter: hypoteser om sporogenesens ursprung vid övergången mellan alger och växter. New Phytol. 190, 875-881. doi: 10.1111/j.1469-8137.2011.03709.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Buendía-Monreal, M., and Gillmor, C. S. (2017). Konvergent repression av miR156 av socker och CDK8-modulen i Arabidopsis Mediator. Dev. Biol. 423, 19-23. doi: 10.1016/j.ydbio.2017.01.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Busch, A., Horn, S., Mühlhausen, A., Mummenhoff, K. och Zachgo, S. (2012). Corolla monosymmetri: evolution av en morfologisk nyhet i Brassicaceae-familjen. Mol. Biol. Evol. 29, 1241-1254. doi: 10.1093/molbev/msr297
PubMed Abstract | CrossRef Full Text | Google Scholar
Cartolano, M., Pieper, B., Lempe, J., Tattersall, A., Huijser, P., Tresch, A., et al. (2015). Heterochrony underpins natural variation in Cardamine hirsuta leaf form. Proc. Natl. Acad. Sci. U.S.A. 112, 10539-10544. doi: 10.1073/pnas.1419791112
PubMed Abstract | CrossRef Full Text | Google Scholar
Chitwood, D. H., Klein, L. L., O’Hanlon, R., Chacko, S., Greg, M., Kitchen, C., et al. (2016). Latenta utvecklings- och evolutionära former inbäddade i vinbladet. New Phytol. 210, 343-355. doi: 10.1111/nph.13754
PubMed Abstract | CrossRef Full Text | Google Scholar
Chuck, G., Cigan, A. M., Saeteurn, K. och Hake, S. (2007). Den heterokroniska majsmutanten corngrass1 beror på överuttryck av ett tandem microRNA. Nat. Genet. 39, 544-549. doi: 10.1038/ng2001
PubMed Abstract | CrossRef Full Text | Google Scholar
Cong, B., Liu, J. och Tanksley, S. D. (2002). Naturliga alleler vid ett kvantitativt egenskapslocus för tomatfruktstorlek skiljer sig åt genom heterokroniska reglerande mutationer. Proc. Natl. Acad. Sci. U.S.A. 99, 13606-13611. doi: 10.1073/pnas.172520999
PubMed Abstract | CrossRef Full Text | Google Scholar
Conway, L. J., and Poethig, R. S. (1997). Mutationer hos Arabidopsis thaliana som omvandlar blad till kotyledoner. Proc. Natl. Acad. Sci. U.S.A. 94, 10209-10214. doi: 10.1073/pnas.94.19.10209
PubMed Abstract | CrossRef Full Text | Google Scholar
Costa, M. M. R., Yang, S., Critchley, J., Feng, X., Wilson, Y., Langlade, N., et al. (2012). Den genetiska grunden för naturlig variation i heteroblasty i Antirrhinum. New Phytol. 196, 1251-1259. doi: 10.1111/j.1469-8137.2012.04347.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Cronk, Q. C. B., Needham, I. och Rudall, P. J. (2015). Evolution of catkins: inflorescence morphology of selected Salicaceae in an evolutionary and developmental context. Front. Plant Sci. 6:1030. doi: 10.3389/fpls.2015.01030
PubMed Abstract | CrossRef Full Text | Google Scholar
Czesnick, H., and Lenhard, M. (2015). Storlekskontroll hos växter – lärdomar från blad och blommor. Cold Spring Harb. Perspect. Biol. 7:a019190. doi: 10.1101/cshperspect.a019190
PubMed Abstract | CrossRef Full Text | Google Scholar
Das Gupta, M., and Nath, U. (2015). Divergens i mönster av bladtillväxtpolaritet är associerad med uttrycksdivergens av miR396. Plant Cell 27, 2785-2799. doi: 10.1105/tpc.15.00196
PubMed Abstract | CrossRef Full Text | Google Scholar
De Beer, G. R. (1951). Embryon och förfäder. Oxford: Clarendon Press.
Google Scholar
Donnelly, P. M., Bonetta, D., Tsukaya, H., Dengler, R. E. och Dengler, N. G. (1999). Cellcykling och cellförstoring i Arabidopsis blad under utveckling. Dev. Biol. 215, 407-419. doi: 10.1006/dbio.1999.9443
PubMed Abstract | CrossRef Full Text | Google Scholar
Efroni, I., Blum, E., Goldshmidt, A. och Eshed, Y. (2008). Ett långvarigt och dynamiskt mognadsschema ligger till grund för Arabidopsis bladutveckling. Plant Cell 20, 2293-2306. doi: 10.1105/tpc.107.057521
PubMed Abstract | CrossRef Full Text | Google Scholar
Fahlgren, N., Montgomery, T. A., Howell, M. D., Allen, E., Dvorak, S. K., Alexander, A. L., et al. (2006). Reglering av AUXIN RESPONSE FACTOR3 med TAS3 ta-siRNA påverkar utvecklingstidpunkten och mönsterbildningen i Arabidopsis. Curr. Biol. 16, 939-944. doi: 10.1016/j.cub.2006.03.065
PubMed Abstract | CrossRef Full Text | Google Scholar
Fernandez, D. E. (1997). Utvecklingsmässiga grunder för homeos i embryon av Brassica napus som gror tidigt: fasförändring vid skottspetsen. Development 124, 1149-1157.
PubMed Abstract | Google Scholar
Friedman, W. E. (1999). Uttryck av cellcykeln i spermier från Arabidopsis: Implikationer för förståelsen av mönster för gametogenes och befruktning hos växter och andra eukaryoter. Development 126, 1065-1075.
PubMed Abstract | Google Scholar
Friedman, W. E., and Carmichael, J. S. (1998). Heterochrony and developmental innovation: evolution of female gametophyte ontogeny in gnetum, a highly Apomorphic seed plant. Evolution 52, 1016-1030. doi: 10.1111/j.1558-5646.1998.tb01830.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Geuten, K., and Coenen, H. (2013). Heterokroniska gener i växternas evolution och utveckling. Front. Plant Sci. 4:381. doi: 10.3389/fpls.2013.00381
PubMed Abstract | CrossRef Full Text | Google Scholar
Gillmor, C. S., Silva-Ortega, C. O., Willmann, M. R., Buendía-Monreal, M., and Poethig, R. S. (2014). Arabidopsis Mediator CDK8-modulgener CCT (MED12) och GCT (MED13) är globala regulatorer av utvecklingsfasövergångar. Development 141, 4580-4589. doi: 10.1242/dev.111229
PubMed Abstract | CrossRef Full Text | Google Scholar
Goebel, K. (1908). Einleitung in die Experimentelle Morphologie der Pflanzen. Leipzig: B.G. Teubner.
Google Scholar
Gould, S. J. (1977). Ontogeni och fylogeni. Cambridge, MA: Harvard University Press.
Google Scholar
Guo, C., Xu, Y., Shi, M., Lai, Y., Wu, X., Wang, H., et al. (2017). Repression av miR156 av miR159 reglerar tidpunkten för övergången från juvenil till vuxen i Arabidopsis. Plant Cell 29, 1293-1304. doi: 10.1105/tpc.16.00975
PubMed Abstract | CrossRef Full Text | Google Scholar
Guo, J. (2015). Mekanismer för evolutionen av komplexa och mångfaldigt utarbetade sepalblad hos Iris identifierade genom jämförande analys av utvecklingssekvenser. Am. J. Bot. 102, 819-832. doi: 10.3732/ajb.1400519
PubMed Abstract | CrossRef Full Text | Google Scholar
Haeckel, E. (1875). Die Gastrula und die Eifurchung der Thiere. Jena. Z. Naturwiss. 9, 402-508.
Google Scholar
Harrison, C. J. (2017). Utveckling och genetik i evolutionen av landväxters kroppsplan. Philos. Trans. R. Soc. B Biol. Sci. 372:20150490. doi: 10.1098/rstb.2015.0490
PubMed Abstract | CrossRef Full Text | Google Scholar
Harrison, C. J., and Morris, J. L. (2018). Ursprunget och den tidiga utvecklingen av kärlväxters skott och blad. Philos. Trans. R. Soc. B Biol. Sci. 373:20160496. doi: 10.1098/rstb.2016.0496
PubMed Abstract | CrossRef Full Text | Google Scholar
He, J., Xu, M., Willmann, M. R., Mccormick, K., Hu, T., Yang, L., et al. (2018). Tröskelberoende repression av SPL-genuttryck av miR156/miR157 kontrollerar vegetativ fasförändring i Arabidopsis thaliana. PLoS Genet. 14:e1007337. doi: 10.1371/journal.pgen.1007337
PubMed Abstract | CrossRef Full Text | Google Scholar
Hudson, C. J., Freeman, J. S., Jones, R. C., Potts, B. M., Wong, M. M. L., Weller, J. L., et al. (2014). Genetisk kontroll av heterokroni i Eucalyptus globulus. G3 4, 1235-1245. doi: 10.1534/g3.114.011916
PubMed Abstract | CrossRef Full Text | Google Scholar
Huijser, P., and Schmid, M. (2011). Kontrollen av utvecklingsfasövergångar i växter. Development 138, 4117-4129. doi: 10.1242/dev.063511
PubMed Abstract | CrossRef Full Text | Google Scholar
Hunter, C., Sun, H., and Poethig, R. S. (2003). Arabidopsis heterokroniska gen ZIPPY är en medlem av ARGONAUTE-familjen. Curr. Biol. 13, 1734-1739. doi: 10.1016/j.cub.2003.09.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Hunter, C., Willmann, M. R., Wu, G., Yoshikawa, M., de la Luz Gutiérrez-Nava, M., and Poethig, R. S. (2006). Transverkande siRNA-medierad repression av ETTIN och ARF4 reglerar heteroblasty i Arabidopsis. Development 133, 2973-2981. doi: 10.1242/dev.02491
PubMed Abstract | CrossRef Full Text | Google Scholar
Jia, X. L., Chen, Y. K., Xu, X. Z., Shen, F., Zheng, Q. B., Du, Z., et al. (2017). miR156 växlar på vegetativ fasförändring under reglering av redoxsignaler i äppelplantor. Sci. Rep. 7, 1-13. doi: 10.1038/s41598-017-14671-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Kenrick, P., and Crane, P. R. (1997). Växternas ursprung och tidiga utveckling på land. Nature 389, 33-39. doi: 10.1038/37918
CrossRef Full Text | Google Scholar
Keyte, A. L., and Smith, K. K. (2014). Heterochroni och utvecklingstidsmekanismer: förändrade ontogenier i evolutionen. Semin. Cell Dev. Biol. 34, 99-107. doi: 10.1016/j.semcdb.2014.06.015
PubMed Abstract | CrossRef Full Text | Google Scholar
Kim, J. Y., Oh, J. E., Noh, Y. S. och Noh, B. (2015). Epigenetisk kontroll av fasövergång från ung till vuxen av Arabidopsis SAGA-liknande komplex. Plant J. 83, 537-545. doi: 10.1111/tpj.12908
PubMed Abstract | CrossRef Full Text | Google Scholar
Kostyun, J. L., Preston, J. C. och Moyle, L. C. (2017). Heterokroniska utvecklingsskiften ligger till grund för den florala mångfalden inom Jaltomata ( Solanaceae ). Evodevo 8:17. doi: 10.1186/s13227-017-0080-z
PubMed Abstract | CrossRef Full Text | Google Scholar
Kyozuka, J., Tokunaga, H. och Yoshida, A. (2014). Kontroll av gräsblomställningens form genom finjustering av meristemets fasförändring. Curr. Opin. Plant Biol. 17, 110-115. doi: 10.1016/j.pbi.2013.11.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Lemmon, Z. H., Park, S. J., Jiang, K., Van Eck, J., Schatz, M. C., and Lippman, Z. B. (2016). Utvecklingen av blomställningens mångfald hos nattskuggor och heterokroni under meristemmognad. Genome Res. 26, 1676-1686. doi: 10.1101/gr.207837.116
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, J., Wang, Z., Hu, Y., Cao, Y. och Ma, L. (2017). Polycombgruppproteinerna RING1A och RING1B reglerar den vegetativa fasövergången i Arabidopsis. Front. Plant Sci. 8:867. doi: 10.3389/fpls.2017.00867
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, P., and Johnston, M. O. (2000). Heterokroni i växtutvecklingsstudier genom det tjugonde århundradet. Bot. Rev. 66, 57-88. doi: 10.1007/BF02857782
CrossRef Full Text | Google Scholar
Li, S., Liu, L., Zhuang, X., Yu, Y., Liu, X., Cui, X., et al. (2013). MikroRNAs hämmar översättningen av mål-mRNAs på det endoplasmatiska retikulumet i Arabidopsis. Cell 153, 562-574. doi: 10.1016/j.cell.2013.04.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Lippman, Z. B., Cohen, O., Alvarez, J. P., Abu-Abied, M., Pekker, I., Paran, I., et al. (2008). Skapandet av en sammansatt blomställning hos tomat och besläktade nattskuggor. PLoS Biol. 6:e288. doi: 10.1371/journal.pbio.0060288
PubMed Abstract | CrossRef Full Text | Google Scholar
Lord, E. M., and Hill, J. P. (1987). ”Evidence for heterochrony in the evolution of plant form,” in Development as an Evolutionary Process, eds R. A. Raff and E. C. Raff (New York, NY: Alan R. Liss), 47-70.
Google Scholar
Maugarny-Calés, A., and Laufs, P. (2018). Att få blad i form: en molekylär, cellulär, miljömässig och evolutionär synvinkel. Development 145:dev161646. doi: 10.1242/dev.161646
PubMed Abstract | CrossRef Full Text | Google Scholar
Meinke, D. W. (1992). En homoetisk mutant av Arabidopsis thaliana med bladiga kotyledoner. Science 258, 1647-1649. doi: 10.1126/science.258.5088.1647
PubMed Abstract | CrossRef Full Text | Google Scholar
Naghiloo, S., and Claßen-Bockhoff, R. (2017). Utvecklingsförändringar i tid och rum främjar evolutionär diversifiering av blommor: en fallstudie i Dipsacoideae. Front. Plant Sci. 8:1665. doi: 10.3389/fpls.2017.01665
PubMed Abstract | CrossRef Full Text | Google Scholar
Nikolov, L. A., Tomlinson, P. B., Manickam, S., Endress, P. K., Kramer, E. M. och Davis, C. C. (2014). Holoparasitiska Rafflesiaceae besitter de mest reducerade endofyterna och ger ändå upphov till världens största blommor. Ann. Bot. 114, 233-242. doi: 10.1093/aob/mcu114
PubMed Abstract | CrossRef Full Text | Google Scholar
Nodine, M. D., and Bartel, D. P. (2010). MicroRNAs förhindrar för tidigt genuttryck och möjliggör mönsterbildning under växtembryogenes. Genes Dev. 24, 2678-2692. doi: 10.1101/gad.1986710
PubMed Abstract | CrossRef Full Text | Google Scholar
Ojeda, D. I., Jaén-Molina, R., Santos-Guerra, A., Caujape-Castells, J. och Cronk, Q. (2017). Temporella, men inte rumsliga, förändringar i uttrycksmönster för gener för kronbladsidentitet är förknippade med förlust av papillerade koniska celler och övergången till fågelpollinering hos makaronesisk lotus (Leguminosae). Plant Biol. 19, 420-427. doi: 10.1111/plb.12551
PubMed Abstract | CrossRef Full Text | Google Scholar
Park, S. J., Jiang, K., Schatz, M. C. och Lippman, Z. B. (2012). Hastigheten för meristemmognad bestämmer blomställningens arkitektur hos tomat. Proc. Natl. Acad. Sci. U.S.A. 109, 639-644. doi: 10.1073/pnas.1114963109
PubMed Abstract | CrossRef Full Text | Google Scholar
Peragine, A., Yoshikawa, M., Wu, G., Albrecht, H. L., and Poethig, R. S. (2004). SGS3 och SGS2/SDE1/RDR6 krävs för juvenil utveckling och produktion av trans-verkande si RNAs i Arabidopsis. Genes Dev. 18, 2368-2379. doi: 10.1101/gad.1231804.nism
CrossRef Full Text | Google Scholar
Pham, T., Day, S. M., Glassford, W. J., Williams, T. M. och Rebeiz, M. (2017). Det evolutionära uppkomsten av ett nytt uttrycksmönster genom ett extremt heterokroniskt skifte. Evol. Dev. 19, 43-55. doi: 10.1111/ede.12215
PubMed Abstract | CrossRef Full Text | Google Scholar
Pires, N. D., and Dolan, L. (2012). Morfologisk evolution hos landväxter: nya mönster med gamla gener. Philos. Trans. R. Soc. B Biol. Sci. 367, 508-518. doi: 10.1098/rstb.2011.0252
PubMed Abstract | CrossRef Full Text | Google Scholar
Poethig, R. S. (2013). Vegetative Phase Change and Shoot Maturation in Plants, 1st Edn. New York, NY: Elsevier Inc. doi: 10.1016/B978-0-12-396968-2.00005-1
PubMed Abstract | CrossRef Full Text | Google Scholar
Porras, R., and Muñoz, J. M. (2000). Cleistogamous capitulum hos Centaurea (Asteraceae): heterokroniskt ursprung. Am. J. Bot. 87, 925-933. doi: 10.2307/2656991
PubMed Abstract | CrossRef Full Text | Google Scholar
Pryer, K. M., and Hearn, D. J. (2009). Evolutionen av bladformen hos marsialiska ormbunkar: bevis för heterokroni. Evolution 63, 498-513. doi: 10.1111/j.1558-5646.2008.00562.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Ronse de Craene, L. (2018). Förståelse av den florala utvecklingens roll i angiosperms blommors evolution: förtydliganden från ett historiskt och fysiskt-dynamiskt perspektiv. J. Plant Res. 131, 367-393. doi: 10.1007/s10265-018-1021-1
PubMed Abstract | CrossRef Full Text | Google Scholar
Rothwell, G. W., Wyatt, S. E. och Tomescu, A. M. F. (2014). Växtutveckling i gränssnittet mellan paleontologi och utvecklingsbiologi: ett organismcentrerat paradigm. Am. J. Bot. 101, 899-913. doi: 10.3732/ajb.1300451
PubMed Abstract | CrossRef Full Text | Google Scholar
Sablowski, R. (2016). Samordning av växtcellers tillväxt och delning: kollektiv kontroll eller ömsesidig överenskommelse? Curr. Opin. Plant Biol. 34, 54-60. doi: 10.1016/j.pbi.2016.09.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Sablowski, R., and Carnier Dornelas, M. (2014). Samspelet mellan celltillväxt och cellcykel i växter. J. Exp. Bot. 65, 2703-2714. doi: 10.1093/jxb/ert354
PubMed Abstract | CrossRef Full Text | Google Scholar
Sharbel, T. F., Voigt, M. L., Corral, J. M., Galla, G., Kumlehn, J., Klukas, C., et al. (2010). Apomictiska och sexuella äggceller av boechera uppvisar heterokroniska globala genuttrycksmönster. Plant Cell 22, 655-671. doi: 10.1105/tpc.109.072223
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, K. K. (2002). Sekvensheterokroni och utvecklingens evolution. J. Morphol. 252, 82-97. doi: 10.1002/jmor.10014
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, K. K. (2003). Time ’ s arrow: heterochrony and the evolution of development. Int. J. Dev. Biol. 47, 613-621.
Google Scholar
Stone, S. L., Braybrook, S. A., Paula, S. L., Kwong, L. W., Meuser, J., Pelletier, J., et al. (2008). Arabidopsis leafy cotyledon2 inducerar mognadsegenskaper och auxinaktivitet: konsekvenser för somatisk embryogenes. Proc. Natl. Acad. Sci. U.S.A. 105, 3151-3156. doi: 10.1073/pnas.0712364105
PubMed Abstract | CrossRef Full Text | Google Scholar
Sun, B., Looi, L. S., Guo, S., He, Z., Gan, E. S., Huang, J., et al. (2014). Tidsmekanism som är beroende av celldelning åberopas av Polycomb eviction i växtstamceller. Science 343:1248559. doi: 10.1126/science.1248559
PubMed Abstract | CrossRef Full Text | Google Scholar
Tian, H. Q., Yuan, T. och Russell, S. D. (2005). Förhållandet mellan dubbel befruktning och cellcykeln i manliga och kvinnliga könsceller av tobak. Sex. Plant Reprod. 17, 243-252. doi: 10.1007/s00497-004-0233-9
CrossRef Full Text | Google Scholar
Tomescu, A. M. F., Wyatt, S. E., Hasebe, M. och Rothwell, G. W. (2014). Tidig evolution av kärlväxternas kroppsplan – de mekanismer som saknas. Curr. Opin. Plant Biol. 17, 126-136. doi: 10.1016/j.pbi.2013.11.016
PubMed Abstract | CrossRef Full Text | Google Scholar
Tsai, A. Y.-L., and Gazzarrini, S. (2014). Trehalos-6-fosfat och SnRK1-kinaser i växters utveckling och signalering: den nya bilden. Front. Plant Sci. 5:119. doi: 10.3389/fpls.2014.00119
PubMed Abstract | CrossRef Full Text | Google Scholar
Vasconcelos, T. N. C., Lucas, E. J., Faria, J. E. Q., and Prenner, G. (2018). Floral heterokroni främjar flexibilitet av reproduktionsstrategier i det morfologiskt homogena släktet Eugenia (Myrtaceae). Ann. Bot. 121, 161-174. doi: 10.1093/aob/mcx142
PubMed Abstract | CrossRef Full Text | Google Scholar
Wahl, V., Ponnu, J., Schlereth, A., Arrivault, S., Langenecker, T., Franke, A., et al. (2013). Reglering av blomning genom trehalose-6-fosfat-signalering i Arabidopsis thaliana. Science 339, 704-707. doi: 10.1126/science.1230406
PubMed Abstract | CrossRef Full Text | Google Scholar
Walcher-Chevillet, C. L., and Kramer, E. M. (2016). Breaking the mold: förståelse av evolutionen och utvecklingen av sidoorgan i olika växtmodeller. Curr. Opin. Genet. Dev. 39, 79-84. doi: 10.1016/j.gde.2016.06.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Wang, J., Czech, B. och Weigel, D. (2009). miR156-regulerade SPL-transkriptionsfaktorer definierar en endogen blomningsväg i Arabidopsis thaliana. Cell 138, 738-749. doi: 10.1016/j.cell.2009.06.014
PubMed Abstract | CrossRef Full Text | Google Scholar
Willmann, M. R., Mehalick, A. J., Packer, R. L. och Jenik, P. D. (2011). MikroRNAs reglerar tidpunkten för embryomognad i Arabidopsis. Plant Physiol. 155, 1871-1884. doi: 10.1104/pp.110.171355
PubMed Abstract | CrossRef Full Text | Google Scholar
Wu, G., Park, M. Y., Conway, S. R., Wang, J., Weigel, D., and Poethig, R. S. (2009). Den sekventiella verkan av miR156 och miR172 reglerar utvecklingstidpunkten i Arabidopsis. Cell 138, 750-759. doi: 10.1016/j.cell.2009.06.031
PubMed Abstract | CrossRef Full Text | Google Scholar
Wu, G., and Poethig, R. S. (2006). Tidsmässig reglering av skottutveckling i Arabidopsis thaliana av miR156 och dess mål SPL3. Development 133, 3539-3547. doi: 10.1242/dev.02521
PubMed Abstract | CrossRef Full Text | Google Scholar
Xu, M., Hu, T., Smith, M. R. och Poethig, R. S. (2016). Epigenetisk reglering av vegetativ fasförändring i Arabidopsis. Plant Cell 28, 28-41. doi: 10.1105/tpc.15.00854
PubMed Abstract | CrossRef Full Text | Google Scholar
Xu, Y., Guo, C., Zhou, B., Li, C., Wang, H., Zheng, B., et al. (2016). Reglering av vegetativ fasförändring av SWI2/SNF2 kromatinremodellerande ATPas BRAHMA. Plant Physiol. 172, 2416-2428. doi: 10.1104/pp.16.01588
PubMed Abstract | CrossRef Full Text | Google Scholar
Xu, M., Leichty, A. R., Hu, T. och Poethig, R. S. (2018). H2A.Z främjar transkriptionen av MIR156A och MIR156C i Arabidopsis genom att underlätta deponering av H3K4me3. Development 145:dev.152868. doi: 10.1242/dev.152868
PubMed Abstract | CrossRef Full Text | Google Scholar
Yamaguchi, N., Huang, J., Xu, Y., Tanoi, K. och Ito, T. (2017). Finjustering av auxinhomeostas styr övergången från underhåll av blommiga stamceller till gynoeciumbildning. Nat. Commun. 8:1125. doi: 10.1038/s41467-017-01252-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Yang, L., Xu, M., Koo, Y., He, J., and Poethig, R. S. (2013). Sugar promotes vegetative phase change in Arabidopsis thaliana by repressing the expression of MIR156A and MIR156C. eLife 2:e00260. doi: 10.7554/eLife.00260.
PubMed Abstract | CrossRef Full Text | Google Scholar
Yu, S., Li, C., Zhou, C. M., Zhang, T. Q., Lian, H., Sun, Y., et al. (2013). Sugar is an endogenous cue for juvenile-to-adult phase transition in plants. eLife 2:e00269. doi: 10.7554/eLife.00269
PubMed Abstract | CrossRef Full Text | Google Scholar
Yu, S., Zhang, Y., Horrer, D., Zhang, T., Hao, Y., Feng, Y., et al. (2012). Gibberellin reglerar Arabidopsis blomövergång genom miR156-märkta SQUAMOSA-promotorbindande – liknande transkriptionsfaktorer. Plant Cell 24, 3320-3332. doi: 10.1105/tpc.112.101014
PubMed Abstract | CrossRef Full Text | Google Scholar
Leave a Reply