Anatomi av hjärtkörtlar och Atrioventrikulärt specialiserat ledningssystem | Revista Española de Cardiología
INLEDNING
De klassiska studierna av Stannius1 från 1852 var de första som föreslog att hjärtats ledningsförmåga var myogenisk. För ungefär hundra år sedan visades att specialiserad muskelvävnad var ansvarig för initieringen och spridningen av hjärtslagen. År 1906 bekräftade Sunao Tawara2 existensen av ett muskulärt buntband som beskrevs av His3 redan 1893. Även 1906 bekräftade Keith och Flack4 existensen av His-Tawara-systemet. Ett år senare beskrev de strukturen hos den sinoatriella (SA) noden.5
Även om Purkinje6 var den förste som beskrev specialiserade ventrikelfibrer var han omedveten om deras betydelse för hjärtats struktur, och det var Tawara2 som visade att det muskelbuntel som His hade beskrivit var kontinuerligt med de ventrikulära Purkinjefibrerna.6 Tawaras studier har nyligen översatts till engelska7 , även om den första översättningen av en del av hans arbete till detta språk gjordes av Robb8 i hans lärobok från 1965. Dessa arbeten är grundläggande läsning för alla forskare som vill studera hjärtats konduktionssystem (CS).
KOMPONENTER AV DET SPECIFIKA ATRIOVENTRIKULA KONDUKTIONSSYSTEMET
Muskelbunten som förbinder förmaken med ventriklarna beskrevs av His3 som en ”penetrerande bunt”. His observerade dock inte den histologiska fortsättningen av detta buntband i höger förmak med den atrioventrikulära (AV) noden, ventriklarna eller de ventrikulära Purkinjecellerna. Det var Tawara2 som upptäckte detta samband när han arbetade för sin doktorsexamen under ledning av Aschoff. Tidigare, 1893, hade både Kent9 och His3 beskrivit muskulära AV-förbindelser som orsakade stor förvirring under många år.10 Senare observerades att dessa förbindelser inte fanns i friska hjärtan, utan i sjuka hjärtan.
Med utgångspunkt i Kents och His upptäckter sökte både kliniska kardiologer och fysiologer efter den struktur som ansvarar för att generera hjärtimpulsen. Man misstänkte att denna var belägen i det område där den övre hålvenen (vena cava superior) och det högra förmaket (atrium) förenas; under experimentella förhållanden är detta den sista delen av hjärtat som slutar slå (det så kallade ultimum moriens). År 1907 urskilde Keith och Flack5 SA- eller sinusknutan hos alla däggdjur som de studerade, inklusive människor. Dess ingående celler ansågs vara platsen för ursprunget av hjärtimpulsen.
SC uppstår i SA-knutan, som finns i det övre främre högra förmaket (figur 1). AV-knutan finns i ett lägre, bakre läge i förmaket. CS sträcker sig från AV-knutan till det penetrerande His-bunten och delar sig sedan i vänster och höger buntgrenar som går ner genom den interventrikulära septum, omslutna av en bindvävsskida som isolerar dem från den omgivande muskelvävnaden. Inne i myokardiet är de kontinuerliga med Purkinje-nätverket (figur 1).
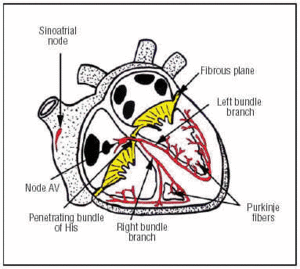
Fig. 1. Schematisk framställning av hjärtats konduktionssystem (rött). Den penetrerande Bundle of His perforerar det fibrösa atrioventrikulära (AV) planet.
Morfologiskt-makroskopiska intresseområden
Flera makroskopiska intresseområden hjälper till att lokalisera hjärtats CS. SA-knutan, som är subepikardiell (figur 2a), är kilad i korsningen mellan muskulaturen i den övre vena cava superior och den i förmaksbilagan. Dess bas ligger mittemot den terminala kammen. Avståndet mellan SA-knutan och epikardiet är 0,3±0,1 mm.11 Hos cirka 10 % av personerna sträcker sig inte knutan mot den nedre vena cava utan ligger i en hästskoform runt den nedre delen av den övre vena cava-öppningen.12 AV-knutan finns vid basen av förmaksseptum vid toppen av ett triangulärt område som först illustrerades av Koch.13 Denna triangel är belägen på endokardialytan av höger förmak (figurerna 2b och c), avgränsas främre av insatsen av trikuspidalklaffens septalblad och bakre av en fibrös sena som kallas Todaros sena. Denna sena är den fibrösa subendokardiella fortsättningen av den Eustachiska klaffen och går in i förmaksmuskulaturen som skiljer sinus coronaris öppning från fossa ovale. Triangelns spets bildas överst av korsningen mellan de främre och bakre gränserna som nämns ovan, vilket motsvarar hjärtats centrala fibrösa kropp (CFB). Triangelns bas utgörs av koronarsinusens mynning tillsammans med höger förmaks vestibulum som stödjer trikuspidalklaffens septalblad. Denna bas är känd för elektrofysiologer som den septala isthmusen, och det är här som radiofrekvensablation av den långsamma banan utförs hos patienter med AV-nodal reentrant takykardi.14
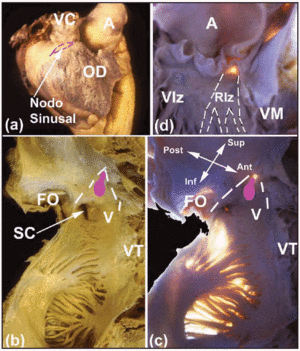
Fig. 2. A: Sidovisning från epikardiet av höger förmak med SA-knutens plats markerad med den streckade rosa linjen. B och C: Endokardiell bild (normal och med transilluminering) av höger förmaks bakre och septala väggar för att visa den ovala fossa (OF) och gränserna för Kochs triangel (streckade vita linjer), Todazos sena (TT) och insättningen av trikuspidalklaffens septala spets (TV). Höger förmaks vestibulum (V) och koronarsinusens mynning (CS) utgör den nedre gränsen. AV-knutans läge visas av en oval knöl (rosa). D: Vänster ventrikelvy för att visa det membranösa septumet (transillumination). Detta är den punkt där His-bunten kommer ut och dess kontinuitet med de högra och vänstra buntgrenarna. Den vänstra buntgrenen (LBB) är markerad med streckade vita linjer. A anger aorta; AA, atrial appendage; VC, superior vena cava; RV, höger ventrikel; LV, vänster ventrikel; MV, mitralisklaff.
Fortsättningen av AV-ledningen sker via det penetrerande His-bunten, den enda delen av ledningsaxeln som perforerar CFB. CFB bildas av föreningen av bindväven i aortaklaffens och mitralisklaffens klaffblad med trikuspidalklaffens septalblad – den så kallade högra fibrösa trigonen – och den membranösa delen av interventrikulära septum. I många däggdjurshjärtan är trigonen fibrös, men nöthjärtan har en central massa av ben eller brosk (os cordis). Däremot är den fibrösa vävnaden i kaskelotet CFB mycket lös. Den membranösa delen eller septum, som kan variera i längd, är en bra vägledning för att lokalisera AV-bunten of His. Detta framträder ovanför denna membranösa del efter att ha korsat den högra fibrösa trigonen (figur 2d) och delar sig sedan i vänster och höger buntgrenar. Den högra grenen passerar genom septummuskulaturen vid basen av den mediala papillarmuskeln i höger kammare. Den blir sedan en tunn sträng som tränger djupt in i den septomarginala trabekulationen eller moderatorbandet som förbinder de mediala och främre papillarmusklerna. Den vänstra grenens ursprung ligger under kommissuren mellan aortaklaffens högra och icke-koronära kusper, och går sedan ner genom subendokardiet i septum interventricularis (figur 2d). Dess väg är ibland synlig på grund av den glänsande fibrösa lamina som omsluter den. Den proximala delen av den vänstra grenen är mycket längre än den högra. Ibland ses en tredje gren som kallas ”dead-end tract ”15 i foster- eller spädbarnshjärtan, och denna fortsätter His-bunten i anterio-superiör riktning mot aortans rot.
Struktur av noderna och det atrioventrikulära ledningssystemet
Studier där de histologiska tekniker som användes liknade dem som användes av Tawara2 och senare arbetare som Davies16 och Truex et al17 (för att nämna några) har visat att CS hos människor är arrangerat på ett sätt som är ganska likt andra däggdjurs (med små variationer mellan arter och mellan hjärtan). Tawara2 rapporterade att de specialiserade myocyterna skiljs från de normala eller arbetande myocyterna genom ett tunt lager bindväv som är synligt i ljusmikroskopet, och på detta bygger de kriterier som Aschoff18 och Mönckeberg19 föreslagit för histologisk identifiering av det specialiserade myokardiet. Enkelt uttryckt kan man säga att specialiserade myocyter skiljer sig från arbetande myocyter när de betraktas i ljusmikroskop och att de kan ”följas” från ett histologiskt snitt till nästa. I sin monografi föredrog Robb8 att definiera den ledande vävnaden med termen ”connecting” snarare än ”conducting” system, eftersom histologiska preparat bättre definierar cellmorfologi än funktion. Han observerade också skillnader i texturen hos det specialiserade myokardiet beroende på hur färskt obduktionsmaterialet var och vilka fixerings- och färgningsmetoder som användes. Tawara2 var medveten om detta och påpekade heterogeniteten i den specialiserade myocytmorfologin även i histologiska snitt av hjärtat. Inom en viss art är de tydligaste skillnaderna relaterade till åldern på de undersökta individerna.20 På senare år har olika molekylära och immunohistokemiska markörer använts för att lokalisera den ledande vävnaden i embryonala hjärtan hos människor och andra däggdjur. Man har dock inte hittat någon specifik markör som kan framhäva denna vävnad hos vuxna människor.
I det normala människohjärtat uppfyller SA- och AV-noderna inte Aschoffs och Mönckebergs kriterier18,19 eftersom de inte är elektriskt isolerade från det omgivande myokardiet genom bindväv eller fettvävnad. De kommer snarare i kontakt med förmaksarbetsfibrerna efter ett litet område som består av övergångsceller. I SA-knutan skiljer Keith och Flack5 mellan sinus- och arbetsceller. Tawara2 angav dock de svårigheter han stötte på när han differentierade AV-knutens celler från cellerna i His-bunten. Han föreslog därför att skillnaden mellan dem var rent anatomisk. På grundval av denna definition kallas den del av CS som är helt omsluten av CFB för penetrerande bunt eller His-bunt (figur 3a). Förmaksdelen från det proximala ledningssystemet till His-bunten kallas AV-knutan (figur 3b). Denna anatomiska distinktion är logisk eftersom isoleringen av det penetrerande His-bunten hindrar den från att komma i direkt kontakt med den elektriska aktiviteten i det afferenta förmaket. All förmaksaktivitet måste därför tidigare ledas genom AV-knutan.
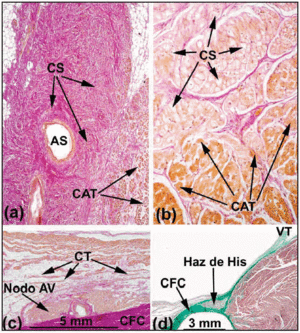
Figur 3. Sagittala histologiska snitt av den sinoatriella (SA) noden i människans (a;x10) och grishjärtat (b;x40) färgade med van Gieson-metoden. Observera kontakten mellan sinuscellerna (SC) och de arbetande förmakscellerna (WAC). Sinuscellerna kännetecknas av att de är tydligare och inbäddade i en större mängd bindväv (röd). c: Van Gieson-färgat snitt av mittzonen i Kochs triangel. Observera formen på den kompakta AV-knutan och övergångscellerna (TC) i kontakt med den kompakta knutens konvexa yta. d: Massons trikromfärgat snitt som visar det penetrerande His-bunten omgivet av fibrös vävnad (grönt) från CFB. SNA anger sinus node artery; CFB, central fibrous body; TV, trikuspidalklaffen.
Den inneboende funktionen hos SA-noden är att vara källan till hjärtimpulsen. SA-knutan hos människor är en välvd eller fusiform struktur. Histologiskt består den av celler som är något mindre än normala arbetsceller och som är ordnade i buntar. Dessa blandas ihop utan någon rumslig ordning, färgar svagt och är inbäddade i en tät bindvävsmatris (figurerna 3 a och 3 b). Med åldern ökar mängden bindväv i förhållande till den yta som upptas av nodalcellerna.21 I knutens periferi blandas specialiserade celler med cellerna från det arbetande myokardiet (figur 3a och b). Dessutom har man beskrivit flera utstrålningar eller förlängningar som interdigiterar med det arbetande förmaksmyokardiet. Dessa tränger intramyokardialt in i terminalkammen och de övre och undre vena cavae. SA-knutan är anordnad runt en artär som kallas sinusnodens artär och som kan löpa centralt eller excentriskt inne i knutan. I 29 procent av människohjärtan förgrenar sig denna artär inuti noden.11 SA-noden är också intimt förknippad med det autonoma nervsystemet. Det har föreslagits att majoriteten av dessa nervfibrer är parasympatiska, medan de sympatiska fibrerna är koncentrerade kring nodens blodkärl.23
AV-knutens inneboende funktion är att fördröja hjärtimpulsen. Hos människor har denna nod en kompakt del och ett område med övergångsceller. Den förstnämnda är halvoval och ligger över CFB (figur 3c). I de delar som ligger nära basen av Kochs triangel delar sig den kompakta delen av noden i två förlängningar eller förlängningar. Den artär som vaskulariserar AV-knutan finns vanligtvis mellan dessa. Längden på dessa förlängningar varierar från ett hjärta till ett annat.24 Storleken på övergångscellerna ligger mellan AV-nodens och förmaksarbetscellernas storlek. De är omgivna av en större mängd bindvävsceller än den som täcker arbetscellerna, men de är inte isolerade från det intilliggande myokardiet. Snarare bildar de ett slags bro mellan det arbetande och nodala myokardiet och samlar in elektrisk information från förmaksväggarna och överför den till AV-knutan.
Kontroverser råder om hur impulsen från SA-knutan når AV-knutan. Vissa författare har föreslagit att det finns specialiserade banor mellan dem.25 Våra studier stöder inte denna idé utan gynnar hypotesen att de arbetande muskelfibrerna i sig (och deras geometriska arrangemang i förmaksväggarna) är ansvariga för att ledningen är snabbare i vissa områden av förmaket än i andra.26
AV-knutan fortsätter distalt med det genomträngande His-bunten (figur 3d), även om det finns små skillnader när det gäller cellulära arrangemanget mellan dessa två strukturer, bland annat är His-bunkerns celler arrangerade på ett mer parallellt sätt. Förklaringen till detta kan vara morfologisk: His-bunten börjar omges av CFB:s bindväv och blir på så sätt en ledningsbana som tar information till ventriklarna.
Hundens AV-nod är mindre än människans, men har en längre penetrerande His-bundel.27 Vissa författare28 tolkar detta som att en del av hundens AV-nod ligger inom CFB. Hos kaninen beskriver andra författare29 en del av His-bunten som om den utgjorde en del av AV-knutan, men detta är ett misstag (figur 4a-d). Den mest framträdande morfologiska skillnaden mellan AV-knutan hos hunden och AV-knutan hos kanin och människa är att den förstnämnda inte är täckt av övergångsceller. Hos råttor (med en vilohjärtfrekvens som är 10 gånger snabbare än hos hundar och människor) är AV-knutan proportionellt sett jämförbar med den hos hundar, men CFB är mindre.
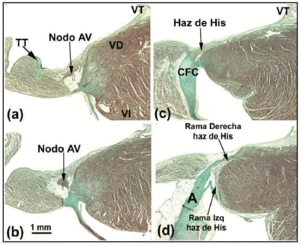
Figur 4. Den här sammansatta figuren visar den atrioventrikulära (AV) noden plus His-bunten och dess höger och vänster buntgrenar hos kaninen. Horisontellt streck i b representerar 1 mm (samma för alla bilder). Massons trichromfärgning. A anger aorta; TT, Todaros sena; RV, höger kammare; LV, vänster kammare; TV, trikuspidalklaff.
När ledningsnätets histologiska bana följs i riktning mot det penetrerande His-bunten, ser man att det senare vänder sig åt vänster i många människohjärtan och kommer ut på den interventrikulära septumets muskulära krön. Omgiven av bindväv från CFB kan längden på His-bunten variera innan den delas upp i den vänstra och den högra buntgrenen. Den förstnämnda grenen kaskaderar över den vänstra sidan av septum interventricularis (figur 5a och c). Delningen av His buntle of His liknar en jockey som sitter på huk över det interventrikulära septumets muskulära krön (figur 5a). Ibland är den dock avvikande åt vänster (figur 5c). När detta inträffar går den högra grenen in i insidan av septummuskulaturen (figur 5b) och uppträder i den högra ventrikeln i samband med insättningen av den mediala papillarmuskeln.
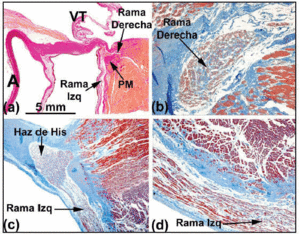
Fig. 5. a: van Gieson-färgat snitt som visar His-bunten (människohjärtat) över membranpartiet (MP) i det interventrikulära septumet. b: Vid sitt ursprung är den högra grenen i detta hjärta intramyokardiell och omges av bindväv (blått) (Jones’ trichromfärgning, x20). c: Delningen av His-bunten i detta hjärta är förskjuten över den vänstra sidan av det interventrikulära septumets muskulära krön och sjunker ner i längsled under vänster kammares endokardium (Jones´ trichrome, x5). d: Lägg märke till den vänstra grenens täckande bindvävshölje (blått) (Jones´ trichrome, x10). A anger aorta; E, endokard; TV, trikuspidalventil.
Längs sina proximala förlopp är höger och vänster buntgrenar täckta av en fibrös lamina (figurerna 5b och d). Som Tawara2 visade (figur 6a) är den vänstra grenen hos människor typiskt uppdelad i tre fascikler med omfattande interkommunikation. Dessa fascikler blir förgrenade i kammarspetsen och sträcker sig till insidan av mitralisklaffens två papillarmuskler, men också tillbaka längs kammarväggarna mot hjärtbasen. Mer distalt, i toppen av ventriklarna i människohjärtat, blir det nästan omöjligt att spåra Purkinjefibrernas förgreningar eftersom dessa förlorar sin fibrösa beläggning och ser ut ungefär som det fungerande myokardiet.
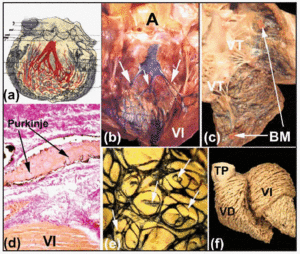
Fig. 6. a: Diagram av Tawara som visar det trifaskulära arrangemanget av den vänstra buntledargrenen hos människan. b och c: Ett färskt kalvhjärta där den högra och vänstra ventrikeln har öppnats. Subendokardiella injektioner av tusch avslöjar de högra och vänstra buntgrenarna och Purkinje-nätverket. Observera i B de tre fasciklerna i den vänstra buntgrenarna (pilar) och i C moderatorbandet (MB). d: Sektion av ett kalvhjärta (van Gieson-färgning, x100) som erhållits efter att ha injicerat bläck i Purkinje-nätverket), som vid sitt ursprung är omslutet av bindväv (rött). e: Purkinje-nätverkets placering i subendokardialt läge i kalvens vänstra kammare. Lägg märke till nätverkets elliptiska arrangemang och utlöpare från kanterna som tränger in i myokardiet (pilar). f: Dissektion av de ventrikulära muskelfibrerna i ett vuxet människohjärta. Observera skillnaden i arrangemang mellan de mediala och djupa lagren i vänster kammare. A anger aorta; PT, lungstam; LV, vänster kammare; LV, vänster ventrikel; TV, trikuspidalklaff.
Subendothelial injektion av bläck är en av de metoder som används för att observera dessa fibrösa skikt och för att påvisa det subendokardiella förloppet av de högra och vänstra buntledargrenarna och deras förgreningar i hovdjurens hjärtan (figurerna 6b och d). Våra studier av får- och kalvhjärtan visar att dessa skiljer sig något från människohjärtan. Kalvhjärtan liknar mer människohjärtan i det avseendet att fasciklerna i den vänstra buntledargrenen vanligtvis är tre till antalet och har sitt ursprung i den övre delen av septum interventricularis (figur 6b). Fårhjärtan uppvisar dock endast två fascikler, och dessa uppträder halvvägs ner i septumväggens längd. I både får- och kalvhjärtan korsar små muskulära trabekler ventrikelhålan – den så kallade ”falska senan” – som inuti dem bär distala förgreningar av His-grenarna mot papillarmusklerna och de intilliggande ventrikelväggarna. På hjärtats högra sida är moderatorbandet i både får- och kalvhjärtat smalare än i människans hjärta, men inuti det finns alltid en utlöpare av höger buntledargren (figur 6c).
I hovdjurshjärtan är det subendokardiella Purkinje-nätverket elliptiskt arrangerat, både i den vänstra och den högra ventrikeln (figur 6e). Från dess kontur uppstår dessutom grenar som penetrerar ventrikelväggarna och leder till nya grenar eller anastamoser med andra grenar (figur 6e). Intramurala grenar av Purkinje-nätverket har dock inte påvisats i människohjärtat.30
En kontroversiell punkt när det gäller Purkinje-nätverket är förekomsten av övergångsceller mellan det fungerande ventrikulära myokardiet och Purkinje-fibrerna.31 De anatomiska och immunohistokemiska studierna av Oosthoek et al30 visar att det i nöthjärtan finns en mycket liten zon med övergångsceller där Purkinje-fibrerna förlorar sin bindvävstäckning. Sådana celler har dock inte observerats i fårhjärtan.32 När Purkinjefibrerna förlorar sitt bindvävshölje går elektriska impulser från CS till de arbetande myocyterna i ventriklarna. Den rumsliga orienteringen av de arbetande myofibrillerna i ventrikelväggarna bestämmer den anisotropa karaktären hos den ventrikulära ledningen (figur 6f).
KONKLUSIONER
Även om det finns skillnader mellan olika arter, liknar strukturen hos knutarna, liksom strukturen hos resten av människans AV-ledningssystem, strukturen hos vanligen använda laboratoriedjur. SA-knutan, den struktur som genererar hjärtimpulsen, är belägen i ena änden av det högra förmaket. Impulserna från den färdas bakåt i förmaksväggarna genom ett invecklat men exakt rumsligt arrangemang av fungerande förmaksfibrer tills de når slutet av förmaket. Där tar AV-nodens övergångsceller emot impulsen och fördröjer den innan den överförs via His-bunten. Det senare korsar det isolerande fibrösa planet mellan förmaken och kamrarna och överför impulsen via två grenar (höger och vänster buntgrenar) till motsvarande kamrar. Var och en av dessa grenar är isolerad av ett sammanbindande hölje av arbetande ventrikulära myocyter. Detta arrangemang tillåter kontakt mellan de specialiserade och arbetande myocyterna endast vid de distala förgreningarna av His-bunten. På detta sätt är AV-ledningssystemet, som till stor del beskrevs av Tawara2 för nästan 100 år sedan, strukturerat för att ge ordning åt överföringen av hjärtimpulser. Kunskap om strukturen och placeringen av specifik ledningsvävnad i hjärtat skulle kunna bidra till att ge lösningar på olika störningar i hjärtrytmen.
Leave a Reply