Anatomia węzłów sercowych i wyspecjalizowanego układu przewodzącego przedsionkowo-komorowego | Revista Española de Cardiología
WSTĘP
Klasyczne badania Stanniusa1 z 1852 roku były pierwszymi, w których zaproponowano, że przewodzenie w sercu jest miogenne. Około sto lat temu wykazano, że wyspecjalizowana tkanka mięśniowa jest odpowiedzialna za inicjację i rozprzestrzenianie się uderzeń serca. W 1906 roku Sunao Tawara2 potwierdził istnienie pęczka mięśniowego opisanego przez Hisa3 już w 1893 roku. Również w 1906 roku Keith i Flack4 potwierdzili istnienie systemu Hisa-Tawary. Rok później opisali oni budowę węzła sino-przedsionkowego (SA).5
Chociaż Purkinje6 jako pierwszy opisał wyspecjalizowane włókna komorowe, nie zdawał sobie sprawy z ich znaczenia w budowie serca, i to właśnie Tawara2 wykazał, że opisana przez Hisa wiązka mięśniowa jest ciągła z komorowymi włóknami Purkinjego.6 Badania Tawary zostały ostatnio przetłumaczone na język angielski,7 chociaż pierwszego przekładu części jego prac na ten język podjął się Robb8 w swoim podręczniku z 1965 roku. Prace te stanowią podstawową lekturę dla wszystkich badaczy, którzy chcieliby studiować układ przewodzący serca (CS).
KOMPONENTY SZCZEGÓLNEGO UKŁADU PRZEWODZENIA ATRIOVENTRICULARNEGO
Pęczek mięśniowy łączący przedsionki z komorami został opisany przez Hisa3 jako „pęczek penetrujący”. Nie zaobserwował on jednak histologicznej kontynuacji tej wiązki w prawym przedsionku z węzłem przedsionkowo-komorowym (AV), komorami lub komórkowymi komórkami Purkinjego. Dopiero Tawara2 rozpoznał to połączenie podczas pracy nad swoim doktoratem pod kierunkiem Aschoffa. Wcześniej, w 1893 roku, zarówno Kent9, jak i His3 opisali mięśniowe połączenia AV, które przez wiele lat były przyczyną wielu nieporozumień.10 Później zaobserwowano, że połączenia te nie występowały w sercach zdrowych, lecz w chorych.
Nawiązując do odkryć Kenta i Hisa, zarówno kardiolodzy kliniczni, jak i fizjolodzy poszukiwali struktury odpowiedzialnej za generowanie impulsu sercowego. Podejrzewano, że znajduje się ona w miejscu połączenia żyły głównej górnej i prawego przedsionka; w warunkach eksperymentalnych jest to ostatnia część serca, która przestaje bić (tzw. ultimum moriens). W 1907 roku Keith i Flack5 wyróżnili węzeł SA lub węzeł zatokowy u wszystkich badanych przez siebie ssaków, w tym u człowieka. Uważano, że komórki wchodzące w jego skład są miejscem pochodzenia impulsu sercowego.
Węzeł CS powstaje w węźle SA, który znajduje się w górnym przednim prawym przedsionku (ryc. 1). Węzeł AV znajduje się w dolnym, tylnym położeniu w przedsionku. CS rozciąga się od węzła AV do penetrującego pęczka Hisa, a następnie dzieli się na lewą i prawą gałąź pęczkową, które zstępują przez przegrodę międzykomorową, otoczone łącznotkankową osłonką izolującą je od otaczającej tkanki mięśniowej. Wewnątrz miokardium są one ciągłe z siecią Purkinjego (ryc. 1).
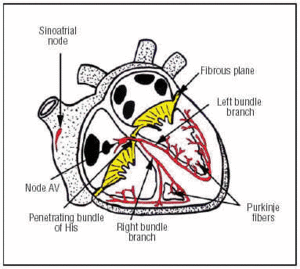
Ryc. 1. Schematyczne przedstawienie układu przewodzącego serca (kolor czerwony). Przenikający pęczek Hisa perforuje włóknistą płaszczyznę przedsionkowo-komorową (AV).
Morfologiczno-makroskopowe obszary zainteresowania
Kilka makroskopowych obszarów zainteresowania pomaga zlokalizować CS serca. Węzeł SA, znajdujący się podwsierdziowo (ryc. 2a), jest zaklinowany w miejscu połączenia mięśnia żyły głównej górnej z mięśniem przedsionka. Jego podstawa znajduje się naprzeciwko grzebienia końcowego. Odległość między węzłem SA a nasierdziem wynosi 0,3±0,1 mm.11 U około 10% osób węzeł nie rozciąga się w kierunku żyły głównej dolnej, lecz leży w kształcie podkowy wokół dolnej części otworu żyły głównej górnej.12 Węzeł AV znajduje się u podstawy przegrody międzyprzedsionkowej, w wierzchołku trójkąta, który po raz pierwszy został przedstawiony przez Kocha.13 Trójkąt ten znajduje się na powierzchni wsierdzia prawego przedsionka (ryc. 2b i c), jest ograniczony z przodu przez wpuklenie płatka przegrodowego zastawki trójdzielnej, a z tyłu przez włókniste ścięgno zwane ścięgnem Todaro. Ścięgno to jest włóknistą podwsierdziową kontynuacją zastawki Eustachiusza i wpukla się do mięśniówki przedsionka, oddzielając otwór zatoki wieńcowej od dołu owalnego. Wierzchołek tego trójkąta tworzy od góry połączenie wyżej wymienionych granic przedniej i tylnej, co odpowiada środkowemu ciału włóknistemu (CFB) serca. Podstawę trójkąta tworzy otwór zatoki wieńcowej wraz z przedsionkiem prawego przedsionka, który wspiera płatek przegrodowy zastawki trójdzielnej. Podstawa ta jest znana elektrofizjologom jako cieśń przegrodowa i to właśnie w tym miejscu wykonuje się ablację drogi wolnej prądem o częstotliwości radiowej u pacjentów z nawrotnym częstoskurczem węzłowym AV.14
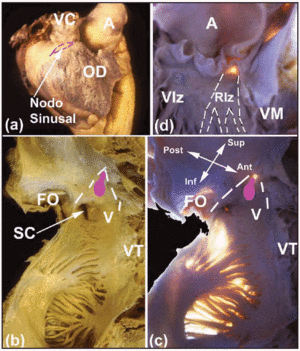
Ryc. 2. A: boczny widok epikardialny prawego przedsionka z miejscem położenia węzła SA zaznaczonym przerywaną różową linią. B i C: widoki wsierdzia (normalny i z transiluminacją) tylnej i przegrodowej ściany prawego przedsionka w celu uwidocznienia dołu owalnego (OF) i granic trójkąta Kocha (przerywane białe linie), ścięgna Todazo (TT) oraz wstawki przegrodowego koniuszka zastawki trójdzielnej (TV). Przedsionek (V) prawego przedsionka i otwór zatoki wieńcowej (CS) tworzą dolną granicę. Lokalizacja węzła AV jest zaznaczona przez owalny guzek (różowy). D: Projekcja lewej komory w celu uwidocznienia przegrody błoniastej (transiluminacja). Jest to miejsce wynicowania pęczka Hisa i jego ciągłości z prawą i lewą odnogą pęczka Hisa. Lewa gałąź pęczkowa (LBB) jest zaznaczona przerywanymi białymi liniami. A oznacza aortę; AA – przedsionek; VC – żyła główna górna; RV – prawa komora; LV – lewa komora; MV – zastawka mitralna.
Dalsza kontynuacja przewodzenia AV odbywa się przez penetrujący pęczek Hisa, jedyną część osi przewodzącej, która perforuje CFB. CFB powstaje w wyniku połączenia tkanki łącznej płatków zastawki aortalnej i mitralnej z płatkiem przegrodowym zastawki trójdzielnej – tzw. prawym trójkątem włóknistym – oraz błoniastą częścią przegrody międzykomorowej. W wielu sercach ssaków trójgraniasty trójkąt jest włóknisty, ale serca bydlęce mają centralną masę kostną lub chrzęstną (os cordis). W przeciwieństwie do tego, tkanka włóknista CFB wieloryba jest bardzo luźna. Część błoniasta lub przegroda, która może mieć różną długość, jest dobrym punktem orientacyjnym do zlokalizowania pęczka AV Hisa. Pojawia się on powyżej części błoniastej po przekroczeniu prawego trójkąta włóknistego (ryc. 2d), a następnie dzieli się na lewą i prawą gałąź pęczka His. Prawa gałąź przechodzi przez mięśniówkę przegrody międzykomorowej u podstawy mięśnia brodawkowatego przyśrodkowego prawej komory. Następnie przechodzi w cienki przewód, który wnika głęboko w trabekulację przegrodowo-marginalną lub pasmo moderatorskie łączące mięsień brodawkowaty przyśrodkowy z mięśniem brodawkowatym przednim. Gałąź lewa ma swój początek poniżej szczeliny między prawym i pozawieńcowym koniuszkiem zastawki aortalnej, a następnie zstępuje przez podwsierdzie przegrody międzykomorowej (ryc. 2d). Jego droga jest niekiedy widoczna dzięki pokrywającej go lśniącej blaszce włóknistej. Proksymalna część gałęzi lewej jest znacznie dłuższa niż prawej. W sercach płodowych i noworodków można niekiedy zaobserwować trzecią gałąź, zwaną „martwym końcem „15 , która kontynuuje wiązkę Hisa w kierunku przednio-przyśrodkowym w kierunku korzenia aorty.
Struktura węzłów i układu przewodzącego przedsionkowo-komorowego
Badania, w których zastosowane techniki histologiczne były podobne do tych stosowanych przez Tawarę2 i późniejszych pracowników, takich jak Davies16 i Truex i wsp.17 (aby wymienić tylko kilku), wykazały, że CS u ludzi jest ułożony w sposób dość podobny do tego, jaki występuje u innych ssaków (z niewielkimi różnicami między gatunkami i między sercami). Tawara2 opisał oddzielenie wyspecjalizowanych miocytów od miocytów prawidłowych lub roboczych cienką warstwą tkanki łącznej widocznej w mikroskopie świetlnym, i na tym opierają się kryteria zaproponowane przez Aschoffa18 i Mönckeberga19 do histologicznej identyfikacji wyspecjalizowanego miokardium. Mówiąc prościej, miocyty wyspecjalizowane odróżniają się od miocytów roboczych w mikroskopie świetlnym i można je „śledzić” z jednego przekroju histologicznego na drugi. W swojej monografii Robb8 wolał określać tkankę przewodzącą terminem „układ łączący”, a nie „przewodzący”, ponieważ preparaty histologiczne lepiej definiują morfologię komórek niż ich funkcję. Zaobserwował on również różnice w strukturze wyspecjalizowanego miokardium w zależności od świeżości materiału autopsyjnego oraz zastosowanych metod utrwalania i barwienia. Tawara2 zdawał sobie z tego sprawę i zwrócił uwagę na heterogenność morfologii wyspecjalizowanych miocytów nawet w wycinkach histologicznych serca. W obrębie danego gatunku najbardziej oczywiste różnice są związane z wiekiem badanych osobników.20 W ostatnich latach do lokalizacji tkanki przewodzącej w embrionalnych sercach ludzi i innych ssaków stosowano różne markery molekularne i immunohistochemiczne. Nie znaleziono jednak specyficznego markera, który mógłby wyróżnić tę tkankę u dorosłych ludzi.
W prawidłowym sercu ludzkim węzły SA i AV nie spełniają kryteriów Aschoffa i Mönckeberga18,19 ponieważ nie są elektrycznie izolowane od otaczającego mięśnia sercowego przez tkankę łączną lub tłuszczową. Wchodzą one raczej w kontakt z włóknami roboczymi przedsionków po niewielkim obszarze zbudowanym z komórek przejściowych. W węźle SA Keith i Flack5 rozróżnili komórki zatokowe i robocze. Tawara2 wskazał natomiast na trudności, jakie napotkał w odróżnieniu komórek węzła AV od komórek pęczka Hisa. Zaproponował zatem, że różnica między nimi ma charakter czysto anatomiczny. Na podstawie tej definicji część CS całkowicie osłonięta przez CFB nazywana jest pęczkiem penetrującym lub pęczkiem Hisa (ryc. 3a). Część przedsionka od proksymalnego układu przewodzącego do pęczka Hisa nazywa się węzłem AV (ryc. 3b). To anatomiczne rozróżnienie jest logiczne, ponieważ izolacja penetrującej wiązki Hisa uniemożliwia jej bezpośredni kontakt z aktywnością elektryczną dośrodkowego przedsionka. Wszelka aktywność przedsionków musi więc być wcześniej kierowana przez węzeł AV.
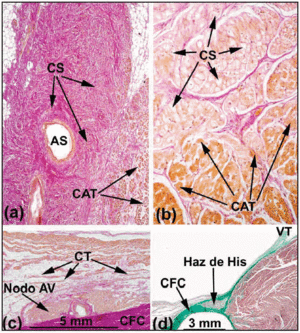
Fig. 3. Sagittal histological sections of the sinoatrial (SA) node of the human (a;x10) and pig heart (b;x40) stained with the van Gieson method. Zwraca uwagę kontakt pomiędzy komórkami zatokowymi (SC) a komórkami pracującego przedsionka (WAC). Komórki zatokowe charakteryzują się większą przejrzystością i są osadzone w większej ilości tkanki łącznej (kolor czerwony). c: Przekrój środkowej strefy trójkąta Kocha wybarwiony metodą van Giesona. Zwraca uwagę kształt zwartego węzła AV oraz komórki przejściowe (TC) stykające się z wypukłą powierzchnią węzła zwartego. d: Wycinek barwiony trichromem Massona ukazujący penetrujący pęczek Hisa otoczony tkanką włóknistą (zielony) z CFB. SNA oznacza tętnicę węzła zatokowego; CFB, centralne ciało włókniste; TV, zastawka trójdzielna.
Wewnętrzną funkcją węzła SA jest bycie źródłem impulsu sercowego. Węzeł SA u człowieka jest strukturą łukowatą lub fuzyjną. Histologicznie składa się on z komórek nieco mniejszych niż normalne komórki robocze, które ułożone są w pęczki. Komórki te mieszają się ze sobą bez ładu przestrzennego, słabo się barwią i są osadzone w gęstej macierzy łącznotkankowej (ryc. 3 a i b). Z wiekiem ilość tkanki łącznej zwiększa się w stosunku do powierzchni zajmowanej przez komórki węzłowe.21 Na obrzeżach węzła wyspecjalizowane komórki mieszają się z komórkami pracującego mięśnia sercowego (ryc. 3a i b). Ponadto opisano liczne promieniowania lub rozszerzenia interdigitalizujące się z pracującym mięśniem przedsionków. Wnikają one wewnątrzsercowo do grzebienia końcowego oraz do żyły głównej górnej i dolnej. Węzeł SA zbudowany jest wokół tętnicy zwanej tętnicą węzłową zatokową, która może przebiegać centralnie lub ekscentrycznie wewnątrz węzła. W 29% ludzkich serc tętnica ta rozgałęzia się wewnątrz węzła.11 Węzeł SA jest również ściśle związany z autonomicznym układem nerwowym. Sugeruje się, że większość tych włókien nerwowych jest przywspółczulna, a włókna współczulne koncentrują się wokół naczyń krwionośnych węzła.23
Nieodłączną funkcją węzła AV jest opóźnianie impulsu sercowego. U człowieka węzeł ten posiada część zwartą oraz obszar komórek przejściowych. Ta pierwsza jest półowalna i leży nad CFB (ryc. 3c). W odcinkach zbliżonych do podstawy trójkąta Kocha część zwarta węzła dzieli się na dwa rozszerzenia lub przedłużenia. Tętnica unaczyniająca węzeł AV znajduje się zwykle pomiędzy nimi. Długość tych rozszerzeń jest różna w różnych sercach.24 Wielkość komórek przejściowych jest pośrednia między komórkami węzła AV a komórkami roboczymi przedsionków. Są one otoczone większą ilością komórek łącznotkankowych niż komórki robocze, ale nie są izolowane od przylegającego mięśnia sercowego. Tworzą one raczej rodzaj pomostu między mięśniem sercowym roboczym a węzłowym i zbierają informacje elektryczne ze ścian przedsionków, przekazując je do węzła AV.
Kontrowersje dotyczą sposobu, w jaki impuls z węzła SA dociera do węzła AV. Niektórzy autorzy sugerują istnienie wyspecjalizowanych szlaków między nimi.25 Nasze badania nie potwierdzają tej koncepcji, ale faworyzują hipotezę, że same pracujące włókna mięśniowe (i ich geometryczne rozmieszczenie w ścianach przedsionków) są odpowiedzialne za to, że przewodzenie jest szybsze w niektórych obszarach przedsionka niż w innych.26
Węzeł AV kontynuuje dystalnie z wnikającym pęczkiem Hisa (ryc. 3d), chociaż istnieją niewielkie różnice pod względem rozmieszczenia komórek między tymi dwiema strukturami, w tym ułożenie komórek pęczka Hisa w sposób bardziej równoległy. Wyjaśnienie tego zjawiska może być morfologiczne: pęczek Hisa zaczyna być otoczony tkanką łączną CFB, stając się w ten sposób przewodnikiem, który przenosi informacje do komór.
Węzeł AV psa jest mniejszy niż u człowieka, ale ma dłuższy penetrujący pęczek Hisa.27 Niektórzy autorzy28 interpretują to w ten sposób, że część węzła AV psa leży w CFB. U królika inni autorzy29 opisują część pęczka Hisa tak, jakby stanowił on część węzła AV, ale jest to błąd (ryc. 4a-d). Najbardziej wyraźną różnicą morfologiczną między węzłem AV psa a węzłami królika i człowieka jest to, że ten pierwszy nie jest pokryty komórkami przejściowymi. U szczurów (u których spoczynkowa czynność serca jest 10 razy szybsza niż u psów czy ludzi) węzeł AV jest proporcjonalnie porównywalny z psim, ale CFB jest mniejszy.
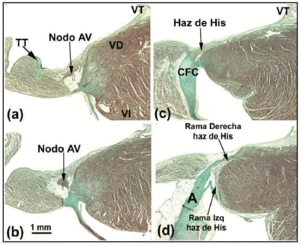
Ryc. 4. Ten złożony rysunek przedstawia węzeł przedsionkowo-komorowy (AV) oraz pęczek Hisa i jego prawe i lewe odgałęzienia pęczka Hisa u królika. Poziomy pasek w b reprezentuje 1 mm (taki sam dla wszystkich obrazów). Barwienie trichromem Massona. A oznacza aortę; TT, ścięgno Todaro; RV, prawa komora; LV, lewa komora; TV, zastawka trójdzielna.
Kiedy podąża się histologiczną trajektorią układu przewodzącego w kierunku penetrującego pęczka Hisa, w wielu ludzkich sercach widać, że skręca on w lewo i wyłania się na grzebieniu mięśniowym przegrody międzykomorowej. Otoczony tkanką łączną pochodzącą z CFB, pęczek Hisa może mieć różną długość, zanim podzieli się na lewą i prawą gałąź pęczka Hisa. Ta pierwsza gałąź biegnie kaskadowo przez lewą stronę przegrody międzykomorowej (ryc. 5a i c). Podział pęczka Hisa przypomina dżokeja przysiadającego nad mięśniowym grzebieniem przegrody międzykomorowej (ryc. 5a). Niekiedy jednak ulega on odchyleniu w kierunku lewym (ryc. 5c). W takim przypadku gałąź prawa wchodzi do wnętrza mięśniówki przegrody (ryc. 5b), pojawiając się w prawej komorze w połączeniu z przyczepem mięśnia brodawkowatego przyśrodkowego.
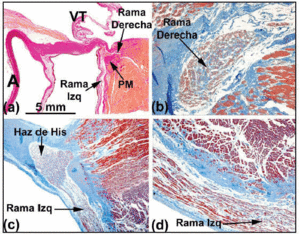
Ryc. 5. a: przekrój barwiony metodą van Giesona przedstawiający pęczek Hisa (serce ludzkie) nad częścią błoniastą (MP) przegrody międzykomorowej. b: gałąź prawa w tym sercu w miejscu swojego początku jest wewnątrzsercowa i otoczona tkanką łączną (kolor niebieski) (barwienie trichromem Jonesa, x20). c: podział pęczka Hisa w tym sercu jest przesunięty na lewą stronę grzebienia mięśniowego przegrody międzykomorowej i zstępuje podłużnie pod wsierdziem lewej komory (trójchrom Jonesa, x5). d: należy zwrócić uwagę na pokrywającą osłonkę łącznotkankową (niebieska) lewej gałęzi (trójchrom Jonesa, x10). A oznacza aortę; E, wsierdzie; TV, zastawka trójdzielna.
Wzdłuż ich proksymalnego przebiegu prawa i lewa odnoga pęczka Hisa pokryte są blaszką włóknistą (ryc. 5b i d). Jak wykazał Tawara2 (ryc. 6a), u ludzi lewa gałąź pęczka Hisa jest zwykle podzielona na trzy powięzi z licznymi połączeniami. Powięzi te stają się rozgałęzione w koniuszku komory i rozciągają się do wnętrza dwóch mięśni brodawkowatych zastawki mitralnej, ale także z powrotem wzdłuż ścian komory w kierunku podstawy serca. Bardziej dystalnie, w koniuszku komór ludzkiego serca, staje się prawie niemożliwe do prześledzenia rozgałęzień włókien Purkinjego, ponieważ tracą one swój włóknisty płaszcz i wyglądają jak pracujące miokardium.
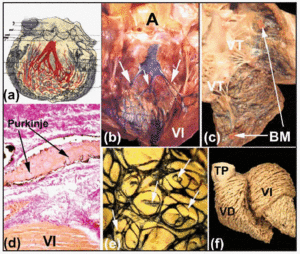
Ryc. 6. a: schemat autorstwa Tawary przedstawiający trójpasmowe ułożenie lewej odnogi pęczka Hisa u ludzi. b i c: świeże serce cielęce, w którym prawa i lewa komora zostały otwarte. Podwsierdziowe iniekcje tuszu indyjskiego uwidaczniają prawe i lewe odgałęzienia pęczkowe oraz sieć Purkinjego. Zauważ w B trzy powięzi lewej odnogi pęczka Hisa (strzałki), a w C pasmo moderatora (MB). d: przekrój serca cielęcia (barwienie van Giesona, x100) uzyskany po wstrzyknięciu tuszu indyjskiego do sieci Purkinjego), która jest otoczona u swego początku tkanką łączną (kolor czerwony). e: podwsierdziowe ułożenie sieci Purkinjego w lewej komorze cielęcia. Zwraca uwagę eliptyczny układ sieci i odrosty od jej brzegów, które wnikają w mięsień sercowy (strzałki). f: przekrój włókien mięśniowych komory dorosłego ludzkiego serca. Należy zwrócić uwagę na różnicę w ułożeniu między warstwą przyśrodkową i głęboką lewej komory. A oznacza aortę; PT, pień płucny; LV, lewa komora; LV, lewa komora; TV, zastawka trójdzielna.
Podśródbłonkowe wstrzyknięcie tuszu indyjskiego jest jedną z metod stosowanych w celu obserwacji tych włóknistych blaszek oraz wykazania podwsierdziowego przebiegu prawej i lewej odnogi pęczka Hisa i ich rozgałęzień w sercach kopytnych (ryc. 6b i d). Nasze badania nad sercami owiec i cieląt wykazały, że różnią się one nieco od serc ludzkich. Serca cieląt są bardziej podobne do serc ludzkich pod tym względem, że powięzi lewej odnogi pęczka Hisa są zazwyczaj trzy i rozpoczynają się w górnej części przegrody międzykomorowej (Ryc. 6b). Natomiast w sercach owczych widoczne są tylko dwie powięzi, które pojawiają się w połowie długości ściany przegrody. W sercu owczym i cielęcym małe trabekule mięśniowe przecinają jamę komory – tzw. fałszywe ścięgno – które wewnątrz przenoszą dystalne odgałęzienia gałęzi Hisa w kierunku mięśni brodawkowatych i przyległych ścian komory. Po prawej stronie serca pasmo moderatorskie zarówno w sercu owcy, jak i cielęcia jest bardziej smukłe niż u człowieka, ale wewnątrz zawsze zawiera odgałęzienie prawej odnogi pęczka Hisa (ryc. 6c).
W sercach kopytnych podwsierdziowa sieć Purkinjego ma układ eliptyczny, zarówno w lewej, jak i w prawej komorze (ryc. 6e). Ponadto z jej obrysu wyrastają odgałęzienia, które penetrują ściany komór, prowadząc do nowych odgałęzień lub zespoleń z innymi odgałęzieniami (ryc. 6e). W sercu ludzkim nie wykazano jednak istnienia śródściennych rozgałęzień sieci Purkinjego.30
Kontrowersyjną kwestią dotyczącą sieci Purkinjego jest istnienie komórek przejściowych między pracującym mięśniem sercowym komory a włóknami Purkinjego.31 Badania anatomiczne i immunohistochemiczne Oosthoeka i wsp.30 wskazują, że w sercach bydlęcych istnieje bardzo mała strefa komórek przejściowych, w której włókna Purkinjego tracą osłonę łącznotkankową. Komórek takich nie zaobserwowano natomiast w sercu owczym.32 Gdy włókna Purkinjego tracą osłonę łącznotkankową, impulsy elektryczne przechodzą z CS do miocytów roboczych komór. Orientacja przestrzenna miofibryli roboczych w ścianach komór decyduje o anizotropowym charakterze przewodzenia komorowego (ryc. 6f).
WNIOSKI
Chociaż istnieją różnice między gatunkami, struktura węzłów, jak również struktura pozostałej części ludzkiego układu przewodzącego AV, jest podobna do powszechnie stosowanej u zwierząt laboratoryjnych. Węzeł SA, struktura generująca impuls sercowy, znajduje się na jednym krańcu prawego przedsionka. Impulsy z niego wędrują w kierunku tylnym w ścianach przedsionków poprzez skomplikowany, ale precyzyjny układ przestrzenny pracujących włókien przedsionkowych aż do końca przedsionka. Na tym końcu komórki przejściowe węzła AV odbierają impuls i opóźniają go przed przekazaniem przez pęczek Hisa. Ten ostatni przecina włóknistą płaszczyznę izolacyjną między przedsionkami a komorami i przekazuje impuls przez dwa odgałęzienia (prawe i lewe odgałęzienie pęczka Hisa) w kierunku odpowiednich komór. Każde z tych odgałęzień jest izolowane przez łączącą je osłonkę z pracujących miocytów komorowych. Taki układ umożliwia kontakt między wyspecjalizowanymi i pracującymi miocytami tylko w dystalnych odgałęzieniach pęczka Hisa. W ten sposób układ przewodzący AV, w dużej mierze opisany przez Tawarę2 prawie 100 lat temu, jest tak skonstruowany, aby nadać porządek przekazywaniu impulsów sercowych. Poznanie struktury i lokalizacji specyficznej tkanki przewodzącej w sercu może pomóc w rozwiązaniu różnych zaburzeń rytmu serca.
Leave a Reply