Frontiers in Plant Science
The Different Types of Heterochrony
In de jaren 1870 identificeerde Ernst Haeckel temporele en ruimtelijke veranderingen in de ontwikkeling van een nakomeling ten opzichte van zijn voorouder als de twee mechanismen die het belangrijkst zijn voor evolutie (Haeckel, 1875). Haeckel noemde ruimtelijke veranderingen heterotopie, en temporele veranderingen heterochronie.
De betekenis van heterochronie is echter veranderd sinds Haeckel voor het eerst de term bedacht. Haeckel gebruikte de term heterochronie om afwijkingen aan te duiden van zijn bekende “Biogenetische Wet”, die stelt dat de opeenvolging van ontwikkelingsgebeurtenissen (ontogenie) grotendeels de opeenvolging van gebeurtenissen in de evolutionaire geschiedenis van de soort (fylogenie) recapituleert (Haeckel, 1875). Met heterochronie werd dus oorspronkelijk een verandering bedoeld in het tijdstip waarop een kenmerk in de ontwikkelingsvolgorde van een organisme verschijnt, ten opzichte van de volgorde in de fylogenie van het organisme. In het midden van de 20e eeuw heeft De Beer (1951) heterochronie losgekoppeld van recapitulatie. Hij gebruikte heterochronie om verschillen aan te duiden in de timing van ontwikkelingsgebeurtenissen bij het vergelijken van twee verwante soorten, om uit te leggen hoe heterochronie diversiteit tussen organismen kon voortbrengen. Gould (1977) bracht het begrip heterochronie opnieuw in verband met recapitulatie, waarbij hij heterochronie definieerde als “veranderingen in het relatieve tijdstip van verschijnen en de ontwikkelingssnelheid van karakters die reeds bij voorouders aanwezig waren”, waarbij hij de nadruk legde op veranderingen in relatieve grootte en vorm, in plaats van in de timing van ontwikkelingsgebeurtenissen, om heterochronie te detecteren. Aan het begin van de 21e eeuw stelde Smith voor dat het nuttiger zou zijn om twee soorten heterochronie te definiëren: “groeiheerochronie,” die de nadruk legt op veranderingen in uiteindelijke grootte en vorm; en “sequentieheerochronie,” die dichter bij het oorspronkelijke gebruik van Haeckel en de Beer staat, en een verklaring van fenotypische variatie mogelijk maakt door veranderingen in de timing van ontwikkelingsgebeurtenissen (Smith, 2002, 2003; Keyte en Smith, 2014).
Sequentie heterochronie (hierna aangeduid als ‘heterochronie’) kan worden ingedeeld in twee categorieën: paedomorfose en peramorfose. In vergelijking met de voorouderlijke ontwikkeling resulteert paedomorfose in een juveniel of eenvoudig resultaat, terwijl peramorfose resulteert in een volwassen of complexer fenotype. Elk van deze twee categorieën van heterochronie kan het gevolg zijn van variatie in de timing van het begin, het einde of de snelheid van een ontwikkelingsproces, zoals voorgesteld door Alberch et al. (1979). Deze variatie kan resulteren in 6 verschillende types van heterochronie (figuur 1). Paedomorfose kan het gevolg zijn van het vroegtijdige einde van een ontwikkelingsproces (progenese), van een vertraagde start van het proces (post-displacement), of van een trager ontwikkelingstempo (neotenie). Peramorfose is het resultaat van een langere ontwikkelingsperiode als gevolg van een later einde (hypermorfose) of een vroeger begin (pre-displacement), of van een hogere ontwikkelingssnelheid (acceleratie) (figuur 1A). Hypothetische voorbeelden voor peramorfose en paedomorfose in de plantenembryogenese en de vegetatieve ontwikkeling zijn weergegeven in de figuren 1B,C.
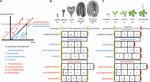
FIGUUR 1. Soorten heterochronie. (A) Schematische voorstelling van de 6 soorten heterochronie. De zwarte lijn geeft de tijd aan die nodig is om een bepaald ontwikkelingsstadium in de voorouderlijke ontogenie te bereiken. Blauwe lijnen tonen de 3 types van paedomorfose: progenese (vroegtijdige aanvang), post-displacement (vertraagde aanvang) en neotenie (tragere ontwikkelingssnelheid). Rode lijnen tonen de 3 soorten peramorfose: hypermorfose (vertraagde offset), pre-displacement (vroegtijdige aanvang), en versnelling (hogere ontwikkelingssnelheid). Tekening in (A) is gebaseerd op Alberch et al. (1979) en Geuten en Coenen (2013). (B) Heterochronie scenario’s voor embryogenese. Arabidopsis embryogenese is hier genomen als de hypothetische voorouderlijke ontwikkeling, voor illustratieve doeleinden verdeeld in vier stadia. Een vijfde stadium, waarin het embryo de eerste twee bladeren plus de twee zaadlobben heeft voortgebracht (aangeduid als embryonale zaailing), wordt voorgesteld als het eindstadium in de peramorfe embryogenese, terwijl de paedomorfe embryogenese naar verwachting zal eindigen in het hartstadium. (C) Heterochronie scenario’s voor vegetatieve ontwikkeling. In dit geval is de vegetatieve ontwikkeling van Arabidopsis, waarbij de plant vóór de bloei cauline/reproductieve bladeren aan de stengel blijft produceren (bovenaan getekend), een hypothetisch geval van peramorfe vegetatieve ontwikkeling, vergeleken met een hypothetische voorouder die in het volwassen stadium bloeit zonder cauline bladeren te produceren. Paedomorfe vegetatieve ontwikkeling leidt volgens de voorspellingen tot bloei in het juveniele stadium.
Hoewel deze classificaties nuttig zijn om veranderingen in ontwikkelingstijdstip te illustreren, is het bij het beschouwen van voorbeelden uit het echte leven vaak moeilijk om onderscheid te maken tussen verschillende soorten heterochronie. In veel gevallen kan evolutie resulteren in verschillende soorten heterochronie, die elk optreden in een discrete fase van een ontwikkelingsproces (besproken in planten door Li en Johnston, 2000).
Heterochronie in de evolutie en diversificatie van planten
Landplanten (embryofyten) hebben veel morfologische innovaties ondergaan sinds hun ontstaan in het midden van het Ordovicium, ongeveer 470 miljoen jaar geleden. Vroege divergerende lineages ontstonden over een periode van meer dan 100 miljoen jaar, tijdens het Siluur en het vroege Devoon (Kenrick en Crane, 1997; Pires en Dolan, 2012; Harrison en Morris, 2018). Embryofyten evolueerden uit een voorouder van zoetwateralgen die verwant was aan de bestaande charofytengroepen Charales, Coleochaetales, en Zygnematales. De overgang naar groei op het land, zoals waargenomen bij bryofyten (hoornmossen, mossen en levermossen), ging gepaard met het ontstaan van sporen, afwisseling van gametofyt- en sporofytgeneraties, eenassige vormen en driedimensionale groei. Innovaties van terrestrische vaatplanten (lycofyten, monilofyten, en spermatofyten) omvatten bifurcatie, onbepaaldheid, sporofytische dominantie, axillaire vertakking, en de vorming van meristemen, bladeren, en wortels (Harrison, 2017).
Verschillende van de bovenstaande evolutionaire stappen zijn in verband gebracht met heterochronie. Voor de oorsprong van sporogenese suggereren gegevens van Bryophyten dat sporen direct uit zygoten werden geproduceerd in een proces met vroegrijpe cytokinese, versnelling van meiose en vertraagde wandafzetting van de zygote naar de mei-osporen (Brown en Lemmon, 2011). Er wordt verondersteld dat de vertakte sporofyt (polisporangiofyt) is geëvolueerd door een langere vegetatieve groei van de apicale cel. Deze langere periode van vegetatieve groei zou resulteren in een verlengde embryonale as, vertakking van de scheut, en een vertraging in de overgang naar reproductieve groei, waarbij het sporangium wordt geproduceerd (Rothwell et al., 2014; Tomescu et al., 2014). Aanvullende studies van plantenfossielen moeten meer bewijs leveren voor oude plantenmorfologieën, waardoor vergelijking tussen hedendaagse en uitgestorven vormen mogelijk wordt (Rothwell et al., 2014).
In extante planten zijn heterochronische veranderingen geïdentificeerd in de gametofytontwikkeling, embryogenese, vegetatieve ontwikkeling, scheutrijping en florale morfogenese. De vrouwelijke gametofyt van Gnetum wijkt structureel af van andere planten door verschillen in de timing van bevruchting en somatische ontwikkeling. Tijdsveranderingen in de voortgang van de celcyclus hebben bijgedragen tot uiteenlopende temporele patronen van spermatogenese en gametenfusie tijdens de bevruchting (Friedman, 1999; Tian et al., 2005). Bij progenese vindt de bevruchting plaats in een vrij kernstadium van de somatische ontwikkeling, een juveniel stadium vergeleken met de voorouderlijke somatische ontogenese, waardoor de differentiatie van eicellen onmogelijk wordt (Friedman en Carmichael, 1998). De apomictische ontwikkeling van Boechera ovules werd geassocieerd met heterochronische genexpressiepatronen in vergelijking met niet-apomictische (seksuele) ovules (Sharbel et al., 2010). De ontwikkeling van Rafflesiaceae, een holoparasitaire plantenfamilie die druivenstokken infecteert, vertoont twee heterochronische verschuivingen: een arrestatie in het proembryonale stadium, wat als een voorbeeld van neotenie kan worden beschouwd, en versnelling van de overgang van de ongedifferentieerde endofyt naar de bloei, waarbij de vegetatieve scheutrijping wordt overgeslagen (Nikolov et al, 2014).
Een Quantitative Trait Loci (QTL) analyse die Eucalyptus globulus populaties met vroegtijdige vegetatieve faseverandering vergelijkt met populaties waarin de vegetatieve faseverandering een aantal jaren wordt uitgesteld, identificeerde de expressie van het microRNA EglMIR156.5 als verantwoordelijk voor de heterochronische variatie in vegetatieve faseverandering in E. globulus (Hudson et al., 2014). Een andere QTL-analyse concludeerde dat heterochronie ten grondslag ligt aan natuurlijke variatie in Cardamine hirsuta bladvorm (Cartolano et al., 2015). QTL-kartering bepaalde dat het effect wordt veroorzaakt door cis-regulerende variatie in de bloemrepressor ChFLC, zodanig dat populaties met laag-expressieve ChFLC-allelen zowel vroege bloei als versnelde verwerving van volwassen bladkenmerken vertonen, met name een verhoogd bladnummer. Morfometrische en QTL-analyses hebben vastgesteld dat heterochronische mutaties bijdragen aan de natuurlijke variatie in Antirrhinum en aan de heteroblastie van de druif (Costa et al., 2012; Chitwood et al., 2016). Een Principale Componenten Analyse (PCA) van de ontogenetische trajecten van bladvorm bij de drie genera van marsileaceeënvarens (Marsilea, Regnellidium, en Pilularia) suggereerde dat ze een paedomorf fenotype vertonen, vergeleken met de complexere voorouderlijke ontwikkeling, veroorzaakt door versnelde groeisnelheid en vroege beëindiging bij een vereenvoudigde bladvorm (Pryer en Hearn, 2009).
Evolutionaire diversiteit in de bloeiwijze-architectuur van de Solanaceae wordt gemoduleerd door heterochronische verschuivingen in de verwerving van het florale lot (Lippman et al, 2008; Park et al., 2012). Een vergelijking van transcriptomen van meristeem rijping van vijf gedomesticeerde en wilde Solanaceae soorten onthulde een piek van expressie divergentie, die lijkt op het “omgekeerde zandloper” model voor dierlijke embryogenese, die stelt dat een midden-ontwikkelingsperiode van divergentie morfologische variatie aandrijft (Lemmon et al., 2016). In grassen resulteert een vertraging in de scheutmeristeem (SM) naar bloemmeristeem (FM) overgang in meer complexe pluimen (Kyozuka et al., 2014). Populieren (Populus sp.) en wilgen (Salix sp.) dragen compacte eenslachtige bloeiwijzen die bekend staan als “katjes”, waarvan is voorgesteld dat ze zijn geëvolueerd uit een vereenvoudiging van de pluimvorm door een vroege SM naar FM overgang (Cronk et al., 2015). Heterochronische veranderingen hebben ook bijgedragen aan natuurlijke variatie in bloeitijd en scheutarchitectuur onder Mimulus guttatus populaties (Baker en Diggle, 2011).
Morfologische diversiteit van de perianth in Dipsacoideae wordt veroorzaakt door heterochronische veranderingen in orgaaninitiatie, specifiek in het aantal kelkbladen (Naghiloo en Claßen-Bockhoff, 2017). De grote vormdiversiteit van kelkbladen onder Iris-soorten is meer te wijten aan heterochronie dan aan heterotopische veranderingen (Guo, 2015). Een studie in Brassicaceae toonde aan dat evolutie van corolla monosymmetrie vanuit de polysymmetrische voorouderlijke bloem gepaard ging met een heterochronische verschuiving in de expressie van CYC2 genen (een clade van TCP transcriptiefactoren) van vroege adaxiale expressie in voorouderlijke bloemmeristemen, naar een latere adaxiale expressie in de ontwikkeling van het kroonblad (Busch et al., 2012). Heterochronische, maar niet heterotope, CYC2-expressie is ook geassocieerd met een verlies van papillate kegelvormige cellen in bloemblaadjes en een verschuiving naar vogelbestuivingssysteem in Lotus (Ojeda et al., 2017). Een paedomorfe morfologie, waarbij de bloemen rijp stuifmeel vasthouden in ongeopende knopachtige structuren, leidde tot gespecialiseerde bestuiving in een clade van Madagaskar wijnstokken (Euphorbiaceae) (Armbruster et al., 2013). De evolutie van cleistogame capitulum vanuit een chasmogame voorouderlijke toestand is een klassiek voorbeeld van paedomorfose, aangezien de cleistogame vorm juveniele kenmerken vertoont (Lord en Hill, 1987). Cleistogamie in Asteraceae is specifiek geëvolueerd door pre-dislocatie en progenese van de bloemontwikkeling, evenals neotenie van alle andere windingen dan het gynoecium (Porras en Muñoz, 2000). De diversiteit van bloemmorfologieën binnen Jaltomata, een Solanaceae genus, is te wijten aan hypermorfose en versnelling van sommige corolla kenmerken (Kostyun et al., 2017). De diversiteit van Azoreaanse vlinderorchideeën wordt ook veroorzaakt door florale heterochronische verschuivingen (Bateman et al., 2014). Onlangs benadrukte Ronse de Craene (2018) het belang van heterochronie in drie ontwikkelingsprocessen: phyllotaxis, de ontwikkeling van gemeenschappelijke meeldraad-petale primordia en obdiplostemonie, waarbij veranderingen in de groeisnelheid in verband worden gebracht met vertraagde orgaaninitiatie. Heterochronische groeisnelheden van de perianth en stijl, en vroege hypanthiumverlenging, zijn verantwoordelijk voor de grote soortendiversiteit binnen het morfologisch homogene Eugenia-geslacht (Vasconcelos et al., 2018). Ten slotte is heterochronische expressie van het fw2.2 allel, dat de celdeling in de vroege vruchtontwikkeling beïnvloedt, verantwoordelijk voor natuurlijke variatie in tomatenvruchtgrootte (Cong et al., 2002).
Studying Heterochronic Mutants to Elucidate Genetic Control of Timing
De studie van mutanten die zijn aangetast in ontwikkelings-timing heeft licht geworpen op genetische paden die morfogenese en ontwikkelingsovergangen controleren. Heterochroniciteit kan veroorzaakt worden door vroegere of latere activatie of repressie van deze pathways.
leafy cotyledon (lec), dicer-like1 (dcl1), en extra cotyledon (xtc) mutanten van Arabidopsis thaliana vertegenwoordigen heterochronische fenotypes die hebben geholpen om zaad rijpingsprogramma’s te definiëren. lec mutanten produceren cotyledons met kenmerken van bladidentiteit (Meinke, 1992), een duidelijk voorbeeld van homeosis: de vervanging van een structuur door een andere. Het is echter vaak moeilijk om homeose en heterochronie van elkaar te onderscheiden, omdat homeose het resultaat kan zijn van zowel heterochronie als heterotopie (Li en Johnston, 2000; Geuten en Coenen, 2013). Tijdens de late embryogenese bevordert LEC2 de zaadrijping en onderdrukt het de postembryonale identiteit (Stone et al., 2008). Net als lec-mutanten vertonen dcl1-mutanten peramorfe fenotypes tijdens de embryogenese, omdat de ontwikkeling van chloroplasten en de expressie van genen voor zaadopslagproteïnen eerder plaatsvindt dan in wild-type embryo’s. DCL1 is nodig voor de biogenese van microRNA’s, die de zaadrijping onderdrukken via de hoofdregulatoren LEC2 en FUSCA3 (Nodine en Bartel, 2010; Willmann et al., 2011). De xtc1, xtc2, en altered meristem programming1 (amp1) mutanten vertonen een homeotisch fenotype waarbij de eerste één of twee bladeren in zaadlobben veranderen (Conway en Poethig, 1997). Bij deze drie mutanten is de overgang van bol naar hart vertraagd, waardoor een vergroot scheutmeristeem ontstaat, wat leidt tot extra orgaanvorming tijdens de embryogenese. Dit fenotype kan worden geïnterpreteerd als hypermorfose, een soort peramorfose, aangezien de embryonale ontwikkeling wordt uitgebreid tot een meer ontwikkelde vorm (zoals het peramorfe scenario in figuur 1B). Indien echter de vegetatieve ontwikkeling als referentiepunt wordt gekozen, zouden lec mutanten een geval van pre-verplaatsing (peramorfose) vertegenwoordigen, en xtc1, xtc2, en amp1 zouden post-verplaatsing (paedomorfose) vertegenwoordigen bij de verwerving van bladidentiteit. AMP1 is nodig voor miRNA-gemedieerde translationele repressie op het ER-membraan (Li et al., 2013).
Genetische regulatie van de overgang van juveniel naar volwassene, ook wel vegetatieve faseverandering genoemd, is bestudeerd met behulp van zowel paedomorfe als peramorfe mutanten. Screeningen voor mutanten met een vroeg-volwassen (peramorf) fenotype leverden allelen op van genen gerelateerd aan kleine RNA biogenese zoals zippy/ago7, sgs3, en rdr6 (Hunter et al., 2003; Peragine et al., 2004), terwijl mutanten met een laat-volwassen (paedomorf) fenotype het gevolg zijn van verhoogde miRNA niveaus (Gillmor et al., 2014; Xu M. et al., 2016, 2018; Guo et al., 2017). De genetische basis voor deze fenotypes wordt verklaard door veranderde expressie van de nauw verwante microRNA’s miR156 en miR157, de belangrijkste regulatoren van vegetatieve faseverandering die werken door SPL-transcriptiefactoren te onderdrukken op zowel transcriptioneel als translationeel niveau, op een drempelafhankelijke manier (Wu en Poethig, 2006; Chuck et al., 2007; Poethig, 2013; He et al., 2018). Zowel MIR156 als SPL genfamilies worden direct gereguleerd door epigenetische markeringen: MIR156 wordt geactiveerd door H3K4me3 en H3K4ac, die worden bevorderd door het SWR1-C complex en de chromatine remodeler BRAHMA, en wordt onderdrukt door H3K27me3, die wordt bevorderd door de Polycomb eiwitten SWINGER en CURLY LEAF en door de chromatine remodeler PICKLE (Xu M. et al., 2016; Xu Y. et al., 2016; Xu M. et al., 2018). SPL-genen worden geactiveerd door histonacetylering gemedieerd door het SAGA-achtig complex en onderdrukt door H2AUb gemedieerd door de Polycomb-eiwitten RING1A en RING1B (Kim et al., 2015; Li et al., 2017). De studie van planten met loss- of gain-of-functie van SPL-genen heeft ook een endogene bloeipadroute gedefinieerd waarin sommige SPL-genen de expressie van miR172 bevorderen, die op zijn beurt de bloei bevordert door de APETALA2-familie bloeirepressoren te onderdrukken (Wang et al., 2009; Wu et al., 2009; Huijser en Schmid, 2011).
Naast belangrijke ontwikkelingsovergangen kan de orgaanmorfogenese ook worden beïnvloed door heterochronische activatie van genetische regulatoren. Bijvoorbeeld, vroege kieming van Brassica rapa embryo’s resulteert in organen met mosaics van cotyledon en blad identiteit (Fernandez, 1997), en differentiële temporele expressie van de klasse II van TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATION CELL FACTOR (TCP) genen resulteert in bladeren met verschillende grootte en vorm (Efroni et al., 2008).
Mechanismen die veranderingen in de timing van de ontwikkeling in planten aansturen (Transcriptionele, Metabole en Cellulaire Heterochronie)
Transcriptionele heterochronie verwijst naar een verandering in de timing van activering of repressie van genexpressie, en wordt vaak veroorzaakt door veranderingen in cis-regulerende genregio’s (Pham et al., 2017). Transcriptionele heterochronie in de hierboven genoemde genetische pathways is een veel voorkomende manier om heterochronische fenotypen te produceren. Metabolische controle van pathways die ontwikkelingsovergangen reguleren (hier aangeduid als “metabole heterochronie”), en temporele controle van celproliferatie, celexpansie en celdifferentiatie (hier aangeduid als “cellulaire heterochronie”) zijn andere mechanismen die heterochronie in planten aansturen (Figuur 2). Transcriptionele, metabolische en cellulaire processen zijn met elkaar verbonden, zodat de moleculaire oorsprong van heterochronie het gevolg kan zijn van een combinatie van mechanismen.

FIGUUR 2. Mechanismen die heterochronie bij planten veroorzaken. (A) Cellulaire heterochronie. De volgorde van de rozetbladeren die tijdens de vegetatieve ontwikkeling worden geproduceerd, wordt getoond. Het fenotype van Arabidopsis Columbia wildtype planten wordt genomen als een hypothetische voorouderlijke toestand; een paedomorf scenario (neotenie) en een peramorf scenario (versnelling) worden ook getoond. Pijlen geven de richting aan van de golf van de celdelingsstilstand in een basipetale gradiënt, waarbij cellen onder de stippellijn zich nog actief delen terwijl de cellen erboven zich uitbreiden en/of differentiëren. (B) Transcriptionele en metabole heterochronie. Een grafische voorstelling van de voorspelde abundantie van het microRNA miR156/7 en zijn targets, de SPL-genen, alsmede de suikerabundantie gedurende de vegetatieve ontwikkeling, corresponderend met de fenotypes in (A). De stippellijnen geven de drempel weer van miR156/7 en SPL abundantie die leidt tot jeugdigheid (boven de drempel) of volwassenheid (onder de drempel). In dit geval resulteert een vertraging in de onderdrukking van miR156 en activering van SPL expressie en suikerproductie in paedomorfose, terwijl een vroegtijdige afbouw van miR156 en vroege activering van SPL expressie en suikerproductie resulteert in peramorfose. Deze tekeningen zijn vereenvoudigde voorstellingen, en geven niet de exacte abundanties in de natuur weer. Het werkelijke patroon van miR156/SPL abundantie is dichter bij dat afgebeeld voor “peramorfose” staat.
Metabole heterochronie kan worden beïnvloed door hormonen, suikers, en redox signalen (Jia et al., 2017). Differentiële biosynthese, transport en perceptie van hormonen zoals auxine, jasmonzuur (JA), gibberelline en abscisinezuur beïnvloeden de heterochronie door regulatoren van ontwikkelingsprocessen aan te sturen. Mutanten in de AUXIN RESPONSE FACTORS (ARF) ARF3 en ARF4 vertragen bijvoorbeeld de volwassen overgang (Fahlgren et al., 2006; Hunter et al., 2006), en auxine homeostase controleert de overgang van bloemstamcelbehoud naar gynoeciumvorming (Yamaguchi et al., 2017). Exogene JA kan de volwassen overgang vertragen door de afname van miR156 expressie uit te stellen (Beydler et al., 2016), en gibberelline versnelt de bloei door SPL genen vrij te maken van repressie door DELLA eiwitten (Yu et al., 2012). De voedingsstatus wordt al sinds het begin van de 20e eeuw in verband gebracht met de controle van vegetatieve faseverandering (Goebel, 1908). Suiker geproduceerd door fotosynthese is noodzakelijk voor de verwerving van volwassen eigenschappen en is gedeeltelijk verantwoordelijk voor de afname van miR156 expressie in de late vegetatieve ontwikkeling (Yang et al., 2013; Yu et al., 2013; Buendía-Monreal en Gillmor, 2017). HEXOKINASE1 (HXK1) en Trehalose-6-fosfaat (T6P) zijn belangrijk voor de suikermedieerde repressie van miR156, waardoor vegetatieve faseverandering en bloei worden bevorderd (Wahl et al., 2013; Yang et al., 2013).
Op cellulair niveau bestaat organogenese uit een opeenvolging van drie fasen: de vestiging van polariteit, celproliferatie, en celexpansie (Walcher-Chevillet en Kramer, 2016). De timing van initiatie en beëindiging van deze stadia is cruciaal voor de grootte en vorm van organen, en heterochronie in deze opeenvolging resulteert in diversificatie van orgaangrootte en -vorm (Figuur 2). De overgang van celproliferatie naar celexpansie en -differentiatie vereist coördinatie tussen de celcyclus en celgroei (Sablowski en Carnier Dornelas, 2014). In bladeren van modelplanten beweegt deze overgang zich als een basipetale golf van celcyclusarrest die begint in het distale deel van het primordium en naar de basis beweegt. Cellen achter het mitotische arrestatiefront raken sterk vacuoleerd en beginnen uit te zetten (Donnelly et al., 1999; Czesnick en Lenhard, 2015). Andere plantensoorten kunnen echter een diffuse groei vertonen, en acropetale of bidirectionele celcyclusarrestgradiënten (Das Gupta and Nath, 2015). De verwerving van fotosynthesecapaciteit is vereist voor de verschuiving van celdeling naar celexpansie (Andriankaja et al., 2012). Deze verschuiving correleert met de rol van suiker in het bevorderen van de Target of Rapamycin (TOR) pathway en het onderdrukken van de Sucrose-non-fermenting1-gerelateerde kinase 1 (SnRK1): TOR en T6P induceren celexpansie door macromoleculaire synthese te bevorderen, terwijl SnRK1 katabolisme bevordert (Tsai en Gazzarrini, 2014; Sablowski, 2016). Twee microRNA’s spelen tegengestelde rollen in deze cellulaire verschuiving: miR319 onderdrukt de expressie van klasse II TCP-factoren, die remmers van celproliferatie zijn, terwijl miR396 de expressie van GROWTH REGULATING FACTORS (GRF’s) beperkt, die de differentiatie vertragen (Das Gupta en Nath, 2015; Maugarny-Calés en Laufs, 2018). De overgang van een onbepaald scheut-apicaal meristeem naar een gedetermineerd bloemmeristeem gaat ook gepaard met temporele regulatie van de cellulaire identiteit. De timing van AGAMOUS activering van KNUCKLES, die op zijn beurt WUSCHEL onderdrukt, bepaalt het temporele venster van onbepaaldheid en bijgevolg de grootte en het aantal organen (Sun et al., 2014).
Conclusie
Een begrip van temporele regulatie van de ontwikkeling van planten is nodig om de diversiteit van plantvormen die we in de natuur zien beter te waarderen, om de morfologische evolutie van planten te verklaren, en om de architectuur van planten te manipuleren ten behoeve van de landbouw. Zoals hierboven geschetst, zijn transcriptionele, metabolische en cellulaire mechanismen, die met elkaar in verband staan, de drijvende kracht achter heterochronie bij bestaande soorten. Verder onderzoek naar deze mechanismen in angiospermen en basale plantensoorten zou meer moeten onthullen over de veranderingen in ontwikkelingstijdstip die de evolutie van de ontwikkeling in planten hebben gestuurd.
Bijdragen van auteurs
MB-M bedacht en schreef het manuscript. CSG bewerkte en reviseerde het manuscript.
Conflict of Interest Statement
De auteurs verklaren dat het onderzoek werd uitgevoerd in afwezigheid van enige commerciële of financiële relaties die zouden kunnen worden opgevat als een potentieel belangenconflict.
Alberch, P., Gould, S., Oster, G., and Wake, D. (1979). Size and shape in ontogeny and phylogeny. Paleobiology 5, 296-317. doi: 10.1017/S0094837300006588
CrossRef Full Text | Google Scholar
Andriankaja, M., Dhondt, S., DeBodt, S., Vanhaeren, H., Coppens, F., DeMilde, L., et al. (2012). Exit van proliferatie tijdens bladontwikkeling in Arabidopsis thaliana: een niet-zo-gradueel proces. Dev. Cell 22, 64-78. doi: 10.1016/j.devcel.2011.11.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Armbruster, W. S., Lee, J., Edwards, M. E., and Baldwin, B. G. (2013). Floral paedomorphy leads to secondary specialization in pollination of Madagascar Dalechampia (Euphorbiaceae). Evolution 67, 1196-1203. doi: 10.1111/evo.12002
PubMed Abstract | CrossRef Full Text | Google Scholar
Baker, R. L., and Diggle, P. K. (2011). Node-specifieke vertakking en heterochronische veranderingen liggen ten grondslag aan populatieniveau verschillen in Mimulus guttatus (Phrymaceae) scheut architectuur. Am. J. Bot. 98, 1924-1934. doi: 10.3732/ajb.1100098
PubMed Abstract | CrossRef Full Text | Google Scholar
Bateman, R. M., Rudall, P. J., Bidartondo, M. I., Cozzolino, S., Tranchida-Lombardo, V., Carine, M. A., et al. (2014). Speciation via floral heterochrony and presumed mycorrhizal host switching of endemic butterfly orchids on the Azorean archipelago. Am. J. Bot. 101, 979-1001. doi: 10.3732/ajb.1300430
PubMed Abstract | CrossRef Full Text | Google Scholar
Beydler, B. D., Osadchuk, K., Cheng, C.-L., Manak, J. R., and Irish, E. E. (2016). De juveniele fase van maïs ziet upregulatie van stress-respons genen en wordt verlengd door exogene JA. Plant Physiol. 171, 2648-2658. doi: 10.1104/pp.15.01707
PubMed Abstract | CrossRef Full Text | Google Scholar
Brown, R. C., and Lemmon, B. E. (2011). Spores before sporophytes: hypothesizing the origin of sporo-genesis at the algal – plant transition. New Phytol. 190, 875-881. doi: 10.1111/j.1469-8137.2011.03709.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Buendía-Monreal, M., and Gillmor, C. S. (2017). Convergente repressie van miR156 door suiker en de CDK8 module van Arabidopsis Mediator. Dev. Biol. 423, 19-23. doi: 10.1016/j.ydbio.2017.01.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Busch, A., Horn, S., Mühlhausen, A., Mummenhoff, K., and Zachgo, S. (2012). Corolla monosymmetrie: evolutie van een morfologische nieuwigheid in de Brassicaceae familie. Mol. Biol. Evol. 29, 1241-1254. doi: 10.1093/molbev/msr297
PubMed Abstract | CrossRef Full Text | Google Scholar
Cartolano, M., Pieper, B., Lempe, J., Tattersall, A., Huijser, P., Tresch, A., et al. (2015). Heterochronie onderbouwt natuurlijke variatie in Cardamine hirsuta bladvorm. Proc. Natl. Acad. Sci. U.S.A. 112, 10539-10544. doi: 10.1073/pnas.1419791112
PubMed Abstract | CrossRef Full Text | Google Scholar
Chitwood, D. H., Klein, L. L., O’Hanlon, R., Chacko, S., Greg, M., Kitchen, C., et al. (2016). Latente ontwikkelings- en evolutionaire vormen ingebed in het wijnstokblad. New Phytol. 210, 343-355. doi: 10.1111/nph.13754
PubMed Abstract | CrossRef Full Text | Google Scholar
Chuck, G., Cigan, A. M., Saeteurn, K., and Hake, S. (2007). De heterochronische maïsmutant corngrass1 is het resultaat van overexpressie van een tandem microRNA. Nat. Genet. 39, 544-549. doi: 10.1038/ng2001
PubMed Abstract | CrossRef Full Text | Google Scholar
Cong, B., Liu, J., and Tanksley, S. D. (2002). Natural alleles at a tomato fruit size quantitative trait locus differ by heterochronic regulatory mutations. Proc. Natl. Acad. Sci. U.S.A. 99, 13606-13611. doi: 10.1073/pnas.172520999
PubMed Abstract | CrossRef Full Text | Google Scholar
Conway, L. J., and Poethig, R. S. (1997). Mutaties van Arabidopsis thaliana die bladeren in cotyledonen veranderen. Proc. Natl. Acad. Sci. U.S.A. 94, 10209-10214. doi: 10.1073/pnas.94.19.10209
PubMed Abstract | CrossRef Full Text | Google Scholar
Costa, M. M. R., Yang, S., Critchley, J., Feng, X., Wilson, Y., Langlade, N., et al. (2012). De genetische basis voor natuurlijke variatie in heteroblastie bij Antirrhinum. New Phytol. 196, 1251-1259. doi: 10.1111/j.1469-8137.2012.04347.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Cronk, Q. C. B., Needham, I., and Rudall, P. J. (2015). Evolution of catkins: inflorescence morphology of selected Salicaceae in an evolutionary and developmental context. Front. Plant Sci. 6:1030. doi: 10.3389/fpls.2015.01030
PubMed Abstract | CrossRef Full Text | Google Scholar
Czesnick, H., and Lenhard, M. (2015). Size control in plants – lessons from leaves and flowers. Cold Spring Harb. Perspect. Biol. 7:a019190. doi: 10.1101/cshperspect.a019190
PubMed Abstract | CrossRef Full Text | Google Scholar
Das Gupta, M., and Nath, U. (2015). Divergentie in patronen van bladgroei polariteit is geassocieerd met de expressie divergentie van miR396. Plant Cell 27, 2785-2799. doi: 10.1105/tpc.15.00196
PubMed Abstract | CrossRef Full Text | Google Scholar
De Beer, G. R. (1951). Embryo’s en voorouders. Oxford: Clarendon Press.
Google Scholar
Donnelly, P. M., Bonetta, D., Tsukaya, H., Dengler, R. E., and Dengler, N. G. (1999). Cell cycling and cell enlargement in developing leaves of Arabidopsis. Dev. Biol. 215, 407-419. doi: 10.1006/dbio.1999.9443
PubMed Abstract | CrossRef Full Text | Google Scholar
Efroni, I., Blum, E., Goldshmidt, A., and Eshed, Y. (2008). Een langdurig en dynamisch rijpingsschema ligt ten grondslag aan de bladontwikkeling van Arabidopsis. Plant Cell 20, 2293-2306. doi: 10.1105/tpc.107.057521
PubMed Abstract | CrossRef Full Text | Google Scholar
Fahlgren, N., Montgomery, T. A., Howell, M. D., Allen, E., Dvorak, S. K., Alexander, A. L., et al. (2006). Regulering van AUXIN RESPONSE FACTOR3 door TAS3 ta-siRNA beïnvloedt de timing en het patroon van de ontwikkeling in Arabidopsis. Curr. Biol. 16, 939-944. doi: 10.1016/j.cub.2006.03.065
PubMed Abstract | CrossRef Full Text | Google Scholar
Fernandez, D. E. (1997). Developmental basis of homeosis in precociously germinating Brassica napus embryos: phase change at the shoot apex. Development 124, 1149-1157.
PubMed Abstract | Google Scholar
Friedman, W. E. (1999). Expressie van de celcyclus in sperma van Arabidopsis: implicaties voor het begrijpen van patronen van gametogenese en bevruchting bij planten en andere eukaryoten. Development 126, 1065-1075.
PubMed Abstract | Google Scholar
Friedman, W. E., and Carmichael, J. S. (1998). Heterochrony and developmental innovation: evolution of female gametophyte ontogeny in gnetum, a highly Apomorphic seed plant. Evolution 52, 1016-1030. doi: 10.1111/j.1558-5646.1998.tb01830.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Geuten, K., and Coenen, H. (2013). Heterochronische genen in plantenevolutie en -ontwikkeling. Front. Plant Sci. 4:381. doi: 10.3389/fpls.2013.00381
PubMed Abstract | CrossRef Full Text | Google Scholar
Gillmor, C. S., Silva-Ortega, C. O., Willmann, M. R., Buendía-Monreal, M., and Poethig, R. S. (2014). De Arabidopsis Mediator CDK8 module genen CCT (MED12) en GCT (MED13) zijn globale regulatoren van ontwikkelingsfasetransities. Development 141, 4580-4589. doi: 10.1242/dev.111229
PubMed Abstract | CrossRef Full Text | Google Scholar
Goebel, K. (1908). Einleitung in die Experimentelle Morphologie der Pflanzen. Leipzig: B.G. Teubner.
Google Scholar
Gould, S. J. (1977). Ontogenie en fylogenie. Cambridge, MA: Harvard University Press.
Google Scholar
Guo, C., Xu, Y., Shi, M., Lai, Y., Wu, X., Wang, H., et al. (2017). Repressie van miR156 door miR159 reguleert de timing van de juveniele-naar-volwassen overgang in Arabidopsis. Plant Cell 29, 1293-1304. doi: 10.1105/tpc.16.00975
PubMed Abstract | CrossRef Full Text | Google Scholar
Guo, J. (2015). Mechanismen voor de evolutie van complexe en divers uitgewerkte kelkbladen in Iris geïdentificeerd door vergelijkende analyse van ontwikkelingssequenties. Am. J. Bot. 102, 819-832. doi: 10.3732/ajb.1400519
PubMed Abstract | CrossRef Full Text | Google Scholar
Haeckel, E. (1875). Die Gastrula und die Eifurchung der Thiere. Jena. Z. Naturwiss. 9, 402-508.
Google Scholar
Harrison, C. J. (2017). Ontwikkeling en genetica in de evolutie van lichaamsplannen van landplanten. Philos. Trans. R. Soc. B Biol. Sci. 372:20150490. doi: 10.1098/rstb.2015.0490
PubMed Abstract | CrossRef Full Text | Google Scholar
Harrison, C. J., and Morris, J. L. (2018). De oorsprong en vroege evolutie van scheuten en bladeren van vasculaire planten. Philos. Trans. R. Soc. B Biol. Sci. 373:20160496. doi: 10.1098/rstb.2016.0496
PubMed Abstract | CrossRef Full Text | Google Scholar
He, J., Xu, M., Willmann, M. R., Mccormick, K., Hu, T., Yang, L., et al. (2018). Drempelwaarde-afhankelijke repressie van SPL genexpressie door miR156/miR157 regelt vegetatieve faseverandering in Arabidopsis thaliana. PLoS Genet. 14:e1007337. doi: 10.1371/journal.pgen.1007337
PubMed Abstract | CrossRef Full Text | Google Scholar
Hudson, C. J., Freeman, J. S., Jones, R. C., Potts, B. M., Wong, M. M. L., Weller, J. L., et al. (2014). Genetische controle van heterochronie in Eucalyptus globulus. G3 4, 1235-1245. doi: 10.1534/g3.114.011916
PubMed Abstract | CrossRef Full Text | Google Scholar
Huijser, P., and Schmid, M. (2011). De controle van ontwikkelingsfase-overgangen in planten. Development 138, 4117-4129. doi: 10.1242/dev.063511
PubMed Abstract | CrossRef Full Text | Google Scholar
Hunter, C., Sun, H., and Poethig, R. S. (2003). Het Arabidopsis heterochronic gen ZIPPY Is een ARGONAUTE familielid. Curr. Biol. 13, 1734-1739. doi: 10.1016/j.cub.2003.09.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Hunter, C., Willmann, M. R., Wu, G., Yoshikawa, M., de la Luz Gutiérrez-Nava, M., and Poethig, R. S. (2006). Trans-acting siRNA-gemedieerde repressie van ETTIN en ARF4 regelt heteroblasty in Arabidopsis. Development 133, 2973-2981. doi: 10.1242/dev.02491
PubMed Abstract | CrossRef Full Text | Google Scholar
Jia, X. L., Chen, Y. K., Xu, X. Z., Shen, F., Zheng, Q. B., Du, Z., et al. (2017). miR156 schakelt vegetatieve faseverandering in onder de regulatie van redoxsignalen in appelzaailingen. Sci. Rep. 7, 1-13. doi: 10.1038/s41598-017-14671-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Kenrick, P., and Crane, P. R. (1997). De oorsprong en vroege evolutie van planten op het land. Nature 389, 33-39. doi: 10.1038/37918
CrossRef Full Text | Google Scholar
Keyte, A. L., and Smith, K. K. (2014). Heterochronie en ontwikkelings-timing mechanismen: veranderende ontogenieën in de evolutie. Semin. Cell Dev. Biol. 34, 99-107. doi: 10.1016/j.semcdb.2014.06.015
PubMed Abstract | CrossRef Full Text | Google Scholar
Kim, J. Y., Oh, J. E., Noh, Y. S., and Noh, B. (2015). Epigenetische controle van juveniele-naar-volwassen faseovergang door het Arabidopsis SAGA-like complex. Plant J. 83, 537-545. doi: 10.1111/tpj.12908
PubMed Abstract | CrossRef Full Text | Google Scholar
Kostyun, J. L., Preston, J. C., and Moyle, L. C. (2017). Heterochronische ontwikkelingsverschuivingen liggen ten grondslag aan florale diversiteit binnen Jaltomata ( Solanaceae ). Evodevo 8:17. doi: 10.1186/s13227-017-0080-z
PubMed Abstract | CrossRef Full Text | Google Scholar
Kyozuka, J., Tokunaga, H., and Yoshida, A. (2014). Controle van gras bloeiwijze vorm door de fijnafstemming van meristeem faseverandering. Curr. Opin. Plant Biol. 17, 110-115. doi: 10.1016/j.pbi.2013.11.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Lemmon, Z. H., Park, S. J., Jiang, K., Van Eck, J., Schatz, M. C., and Lippman, Z. B. (2016). De evolutie van bloeiwijze diversiteit in de nachtschades en heterochronie tijdens meristeem rijping. Genome Res. 26, 1676-1686. doi: 10.1101/gr.207837.116
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, J., Wang, Z., Hu, Y., Cao, Y., and Ma, L. (2017). Polycomb groep eiwitten RING1A en RING1B reguleren de vegetatieve faseovergang in Arabidopsis. Front. Plant Sci. 8:867. doi: 10.3389/fpls.2017.00867
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, P., and Johnston, M. O. (2000). Heterochronie in plant evolutionaire studies door de twintigste eeuw. Bot. Rev. 66, 57-88. doi: 10.1007/BF02857782
CrossRef Full Text | Google Scholar
Li, S., Liu, L., Zhuang, X., Yu, Y., Liu, X., Cui, X., et al. (2013). MicroRNAs remmen de translatie van doel-mRNAs op het endoplasmatisch reticulum in Arabidopsis. Cell 153, 562-574. doi: 10.1016/j.cell.2013.04.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Lippman, Z. B., Cohen, O., Alvarez, J. P., Abu-Abied, M., Pekker, I., Paran, I., et al. (2008). Het ontstaan van een samengestelde bloeiwijze in tomaat en verwante nachtschades. PLoS Biol. 6:e288. doi: 10.1371/journal.pbio.0060288
PubMed Abstract | CrossRef Full Text | Google Scholar
Lord, E. M., and Hill, J. P. (1987). “Evidence for heterochrony in the evolution of plant form,” in Development as an Evolutionary Process, eds R. A. Raff and E. C. Raff (New York, NY: Alan R. Liss), 47-70.
Google Scholar
Maugarny-Calés, A., and Laufs, P. (2018). Het in vorm krijgen van bladeren: een moleculaire, cellulaire, omgevings- en evolutionaire kijk. Development 145:dev161646. doi: 10.1242/dev.161646
PubMed Abstract | CrossRef Full Text | Google Scholar
Meinke, D. W. (1992). Een homoetische mutant van Arabidopsis thaliana met bladhoudende cotyledonen. Science 258, 1647-1649. doi: 10.1126/science.258.5088.1647
PubMed Abstract | CrossRef Full Text | Google Scholar
Naghiloo, S., and Claßen-Bockhoff, R. (2017). Ontwikkelingsveranderingen in tijd en ruimte bevorderen evolutionaire diversificatie van bloemen: een case study in Dipsacoideae. Front. Plant Sci. 8:1665. doi: 10.3389/fpls.2017.01665
PubMed Abstract | CrossRef Full Text | Google Scholar
Nikolov, L. A., Tomlinson, P. B., Manickam, S., Endress, P. K., Kramer, E. M., and Davis, C. C. (2014). Holoparasitic Rafflesiaceae possess the most reduced endophytes and yet give rise to the world’s largest flowers. Ann. Bot. 114, 233-242. doi: 10.1093/aob/mcu114
PubMed Abstract | CrossRef Full Text | Google Scholar
Nodine, M. D., and Bartel, D. P. (2010). MicroRNAs voorkomen vroegtijdige genexpressie en maken patroonvorming mogelijk tijdens plantenembryogenese. Genes Dev. 24, 2678-2692. doi: 10.1101/gad.1986710
PubMed Abstract | CrossRef Full Text | Google Scholar
Ojeda, D. I., Jaén-Molina, R., Santos-Guerra, A., Caujape-Castells, J., and Cronk, Q. (2017). Temporele, maar niet ruimtelijke, veranderingen in expressiepatronen van petale identiteitsgenen zijn geassocieerd met verlies van papillate conische cellen en de verschuiving naar vogelbestuiving in Macaronesische Lotus (Leguminosae). Plant Biol. 19, 420-427. doi: 10.1111/plb.12551
PubMed Abstract | CrossRef Full Text | Google Scholar
Park, S. J., Jiang, K., Schatz, M. C., and Lippman, Z. B. (2012). De snelheid van meristeemrijping bepaalt de bloeiwijze-architectuur in tomaat. Proc. Natl. Acad. Sci. U.S.A. 109, 639-644. doi: 10.1073/pnas.1114963109
PubMed Abstract | CrossRef Full Text | Google Scholar
Peragine, A., Yoshikawa, M., Wu, G., Albrecht, H. L., and Poethig, R. S. (2004). SGS3 en SGS2/SDE1/RDR6 zijn nodig voor de ontwikkeling van juvenielen en de productie van trans-werkende si RNAs in Arabidopsis. Genes Dev. 18, 2368-2379. doi: 10.1101/gad.1231804.nism
CrossRef Full Text | Google Scholar
Pham, T., Day, S. M., Glassford, W. J., Williams, T. M., and Rebeiz, M. (2017). De evolutionaire oorsprong van een nieuw expressiepatroon door een extreme heterochronische verschuiving. Evol. Dev. 19, 43-55. doi: 10.1111/ede.12215
PubMed Abstract | CrossRef Full Text | Google Scholar
Pires, N. D., and Dolan, L. (2012). Morfologische evolutie in landplanten: nieuwe ontwerpen met oude genen. Philos. Trans. R. Soc. B Biol. Sci. 367, 508-518. doi: 10.1098/rstb.2011.0252
PubMed Abstract | CrossRef Full Text | Google Scholar
Poethig, R. S. (2013). Vegetative Phase Change and Shoot Maturation in Plants, 1st Edn. New York, NY: Elsevier Inc. doi: 10.1016/B978-0-12-396968-2.00005-1
PubMed Abstract | CrossRef Full Text | Google Scholar
Porras, R., and Muñoz, J. M. (2000). Cleistogame capitulum bij Centaurea (Asteraceae): heterochronische oorsprong. Am. J. Bot. 87, 925-933. doi: 10.2307/2656991
PubMed Abstract | CrossRef Full Text | Google Scholar
Pryer, K. M., and Hearn, D. J. (2009). Evolution of leaf form in marsileaceous ferns: evidence for heterochrony. Evolution 63, 498-513. doi: 10.1111/j.1558-5646.2008.00562.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Ronse de Craene, L. (2018). Inzicht in de rol van bloemontwikkeling in de evolutie van angiosperm bloemen: verduidelijkingen vanuit een historisch en fysisch-dynamisch perspectief. J. Plant Res. 131, 367-393. doi: 10.1007/s10265-018-1021-1
PubMed Abstract | CrossRef Full Text | Google Scholar
Rothwell, G. W., Wyatt, S. E., and Tomescu, A. M. F. (2014). Plant evolution at the interface of paleontology and developmental biology: an organism-centered paradigm. Am. J. Bot. 101, 899-913. doi: 10.3732/ajb.1300451
PubMed Abstract | CrossRef Full Text | Google Scholar
Sablowski, R. (2016). Coördinatie van plantencelgroei en -deling: collectieve controle of wederzijdse overeenkomst? Curr. Opin. Plant Biol. 34, 54-60. doi: 10.1016/j.pbi.2016.09.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Sablowski, R., and Carnier Dornelas, M. (2014). Wisselwerking tussen celgroei en celcyclus in planten. J. Exp. Bot. 65, 2703-2714. doi: 10.1093/jxb/ert354
PubMed Abstract | CrossRef Full Text | Google Scholar
Sharbel, T. F., Voigt, M. L., Corral, J. M., Galla, G., Kumlehn, J., Klukas, C., et al. (2010). Apomictische en seksuele ovules van boechera vertonen heterochronische globale genexpressiepatronen. Plant Cell 22, 655-671. doi: 10.1105/tpc.109.072223
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, K. K. (2002). Sequence heterochrony and the evolution of development. J. Morphol. 252, 82-97. doi: 10.1002/jmor.10014
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, K. K. (2003). Time ‘ s arrow: heterochronie en de evolutie van ontwikkeling. Int. J. Dev. Biol. 47, 613-621.
Google Scholar
Stone, S. L., Braybrook, S. A., Paula, S. L., Kwong, L. W., Meuser, J., Pelletier, J., et al. (2008). Arabidopsis leafy cotyledon2 induceert rijpingskenmerken en auxine activiteit: implicaties voor somatische embryogenese. Proc. Natl. Acad. Sci. U.S.A. 105, 3151-3156. doi: 10.1073/pnas.0712364105
PubMed Abstract | CrossRef Full Text | Google Scholar
Sun, B., Looi, L. S., Guo, S., He, Z., Gan, E. S., Huang, J., et al. (2014). Timing mechanisme afhankelijk van celdeling wordt opgeroepen door Polycomb evictie in plantaardige stamcellen. Science 343:1248559. doi: 10.1126/science.1248559
PubMed Abstract | CrossRef Full Text | Google Scholar
Tian, H. Q., Yuan, T., and Russell, S. D. (2005). Verband tussen dubbele bevruchting en de celcyclus in mannelijke en vrouwelijke gameten van tabak. Sex. Plant Reprod. 17, 243-252. doi: 10.1007/s00497-004-0233-9
CrossRef Full Text | Google Scholar
Tomescu, A. M. F., Wyatt, S. E., Hasebe, M., and Rothwell, G. W. (2014). Vroege evolutie van het vasculaire plantenlichaamsplan – de ontbrekende mechanismen. Curr. Opin. Plant Biol. 17, 126-136. doi: 10.1016/j.pbi.2013.11.016
PubMed Abstract | CrossRef Full Text | Google Scholar
Tsai, A. Y.-L., and Gazzarrini, S. (2014). Trehalose-6-fosfaat en SnRK1 kinases in plant ontwikkeling en signalering: het opkomende beeld. Front. Plant Sci. 5:119. doi: 10.3389/fpls.2014.00119
PubMed Abstract | CrossRef Full Text | Google Scholar
Vasconcelos, T. N. C., Lucas, E. J., Faria, J. E. Q., and Prenner, G. (2018). Florale heterochronie bevordert flexibiliteit van reproductieve strategieën in het morfologisch homogene genus Eugenia (Myrtaceae). Ann. Bot. 121, 161-174. doi: 10.1093/aob/mcx142
PubMed Abstract | CrossRef Full Text | Google Scholar
Wahl, V., Ponnu, J., Schlereth, A., Arrivault, S., Langenecker, T., Franke, A., et al. (2013). Regeling van de bloei door trehalose-6-fosfaat signalering in Arabidopsis thaliana. Science 339, 704-707. doi: 10.1126/science.1230406
PubMed Abstract | CrossRef Full Text | Google Scholar
Walcher-Chevillet, C. L., and Kramer, E. M. (2016). Breaking the mold: understanding the evolution and development of lateral organs in diverse plant models. Curr. Opin. Genet. Dev. 39, 79-84. doi: 10.1016/j.gde.2016.06.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Wang, J., Czech, B., and Weigel, D. (2009). miR156-regulerende SPL-transcriptiefactoren definiëren een endogene bloeipadway in Arabidopsis thaliana. Cell 138, 738-749. doi: 10.1016/j.cell.2009.06.014
PubMed Abstract | CrossRef Full Text | Google Scholar
Willmann, M. R., Mehalick, A. J., Packer, R. L., and Jenik, P. D. (2011). MicroRNAs reguleren de timing van de rijping van embryo’s in Arabidopsis. Plant Physiol. 155, 1871-1884. doi: 10.1104/pp.110.171355
PubMed Abstract | CrossRef Full Text | Google Scholar
Wu, G., Park, M. Y., Conway, S. R., Wang, J., Weigel, D., and Poethig, R. S. (2009). De opeenvolgende actie van miR156 en miR172 regelt de timing van de ontwikkeling in Arabidopsis. Cell 138, 750-759. doi: 10.1016/j.cell.2009.06.031
PubMed Abstract | CrossRef Full Text | Google Scholar
Wu, G., and Poethig, R. S. (2006). Temporal regulation of shoot development in Arabidopsis thaliana by miR156 and its target SPL3. Development 133, 3539-3547. doi: 10.1242/dev.02521
PubMed Abstract | CrossRef Full Text | Google Scholar
Xu, M., Hu, T., Smith, M. R., and Poethig, R. S. (2016). Epigenetische regulatie van vegetatieve faseverandering in Arabidopsis. Plant Cell 28, 28-41. doi: 10.1105/tpc.15.00854
PubMed Abstract | CrossRef Full Text | Google Scholar
Xu, Y., Guo, C., Zhou, B., Li, C., Wang, H., Zheng, B., et al. (2016). Regeling van vegetatieve faseverandering door SWI2/SNF2 chromatine remodeling ATPase BRAHMA. Plant Physiol. 172, 2416-2428. doi: 10.1104/pp.16.01588
PubMed Abstract | CrossRef Full Text | Google Scholar
Xu, M., Leichty, A. R., Hu, T., and Poethig, R. S. (2018). H2A.Z bevordert de transcriptie van MIR156A en MIR156C in Arabidopsis door de depositie van H3K4me3 te vergemakkelijken. Development 145:dev.152868. doi: 10.1242/dev.152868
PubMed Abstract | CrossRef Full Text | Google Scholar
Yamaguchi, N., Huang, J., Xu, Y., Tanoi, K., and Ito, T. (2017). Fine-tuning of auxin homeostasis governs the transition from floral stem cell maintenance to gynoecium formation. Nat. Commun. 8:1125. doi: 10.1038/s41467-017-01252-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Yang, L., Xu, M., Koo, Y., He, J., and Poethig, R. S. (2013). Suiker bevordert vegetatieve faseverandering in Arabidopsis thaliana door onderdrukking van de expressie van MIR156A en MIR156C. eLife 2:e00260. doi: 10.7554/eLife.00260
PubMed Abstract | CrossRef Full Text | Google Scholar
Yu, S., Li, C., Zhou, C. M., Zhang, T. Q., Lian, H., Sun, Y., et al. (2013). Sugar is an endogenous cue for juvenile-to-adult phase transition in plants. eLife 2:e00269. doi: 10.7554/eLife.00269
PubMed Abstract | CrossRef Full Text | Google Scholar
Yu, S., Zhang, Y., Horrer, D., Zhang, T., Hao, Y., Feng, Y., et al. (2012). Gibberelline reguleert de bloeiwijze van Arabidopsis via miR156-getargette SQUAMOSA promoter binding – zoals transcriptiefactoren. Plant Cell 24, 3320-3332. doi: 10.1105/tpc.112.101014
PubMed Abstract | CrossRef Full Text | Google Scholar
Leave a Reply