Anatomy of Cardiac Nodes and Atrioventricular Specialized Conduction System | Revista Española de Cardiología
INTRODUCTION
De klassieke studies van Stannius1 in 1852 waren de eerste die stelden dat de hartgeleiding myogeen was. Ongeveer een eeuw geleden werd aangetoond dat gespecialiseerd spierweefsel verantwoordelijk was voor de initiatie en verspreiding van de hartslag. In 1906 bevestigde Sunao Tawara2 het bestaan van een spierbundel die reeds in 1893 door His3 was beschreven. Eveneens in 1906 bevestigden Keith en Flack4 het bestaan van het His-Tawara systeem. Een jaar later beschreven zij de structuur van de sinoatriale knoop (SA).5 Hoewel Purkinje6 de eerste was die gespecialiseerde ventriculaire vezels beschreef, was hij zich niet bewust van hun belang in de structuur van het hart, en het was Tawara2 die aantoonde dat de spierbundel beschreven door His doorlopend was met de ventriculaire Purkinje vezels.6 De studies van Tawara zijn onlangs in het Engels vertaald,7 hoewel de eerste vertaling van een deel van zijn werk in deze taal werd ondernomen door Robb8 in zijn leerboek uit 1965. Deze werken zijn basis lectuur voor alle onderzoekers die het cardiale geleidingssysteem (CS) willen bestuderen.
COMPONENTEN VAN HET SPECIFIEKE ATRIOVENTRICULAIRE GELEIDSSYSTEEM
De spierbundel die de atria met de ventrikels verbindt, werd door His3 beschreven als een “penetrerende bundel”. His heeft echter niet de histologische voortzetting van deze bundel in de rechterboezem met de atrioventriculaire (AV) knoop, de ventrikels, of de ventriculaire Purkinje cellen waargenomen. Het was Tawara2 die deze verbinding herkende toen hij onder leiding van Aschoff aan zijn doctoraat werkte. Eerder, in 1893, hadden zowel Kent9 als His3 gespierde AV-verbindingen beschreven die jarenlang voor veel verwarring zorgden.10 Later werd vastgesteld dat deze verbindingen niet in gezonde harten werden aangetroffen, maar in zieke harten.
In het verlengde van de bevindingen van Kent en His zochten zowel klinisch cardiologen als fysiologen naar de structuur die verantwoordelijk was voor het opwekken van de hartimpuls. Men vermoedde dat deze zich bevond in het gebied waar de superieure vena cava en de rechterboezem samenkwamen; onder experimentele omstandigheden is dit het laatste deel van het hart dat stopt met kloppen (het zogenaamde ultimum moriens). In 1907 onderscheidden Keith en Flack5 de SA- of sinusknoop bij alle zoogdieren die zij bestudeerden, met inbegrip van de mens. De samenstellende cellen ervan werden verondersteld de plaats van oorsprong van de hartimpuls te zijn.
De sinusknoop ontstaat in de SA-knoop, die zich in de bovenste voorste rechterboezem bevindt (figuur 1). De AV-knoop bevindt zich lager, achteraan in de boezem. De CS strekt zich uit van de AV-knoop tot de doordringende bundel van His en splitst zich vervolgens in de linker- en rechterbundeltak die door het interventriculaire septum naar beneden gaan, omhuld door een bindweefselschede die hen isoleert van het omringende spierweefsel. Binnen het myocard lopen zij door in het Purkinje-netwerk (figuur 1).
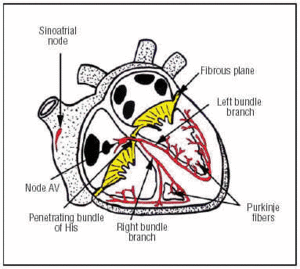
Fig. 1. Diagrammatische weergave van het geleidingssysteem van het hart (rood). De doordringende Bundel van Zijn perforeert het vezelige atrioventriculaire (AV) vlak.
Morfologisch-macroscopische aandachtsgebieden
Verscheidene macroscopische aandachtsgebieden helpen om het cardiale CS te lokaliseren. De SA-knoop, die sub-epicardiale (figuur 2a), is ingeklemd in het kruispunt tussen de musculatuur van de superieure vena cava en die van de atriale appendage. De basis ligt tegenover de terminale kam. De afstand tussen de SA-knoop en het epicard is 0,3±0,1 mm.11 Bij ongeveer 10% van de personen strekt de knoop zich niet uit naar de vena cava inferior, maar ligt hij in een hoefijzervorm rond het onderste deel van de orifice van de vena cava superior.12 De AV-knoop bevindt zich aan de basis van het atriumseptum op de top van een driehoekig gebied dat voor het eerst werd geïllustreerd door Koch.13 Deze driehoek bevindt zich op het endocardiale oppervlak van de rechterboezem (Figuren 2b en c), wordt anterior begrensd door de insertie van de septale leaflet van de tricuspidalisklep, en posterior door een vezelige pees bekend als de pees van Todaro. Deze pees is de vezelige subendocardiale voortzetting van de Eustachiusklep, en gaat over in de atriale musculatuur die de opening van de coronaire sinus scheidt van de fossa ovale. De top van deze driehoek wordt in superieure richting gevormd door de kruising van de bovengenoemde voorste en achterste grens, die overeenkomt met het centrale vezellichaam (CFB) van het hart. De basis van de driehoek wordt gevormd door de opening van de coronaire sinus samen met het voorportaal van de rechterboezem dat het septale blad van de tricuspidalisklep ondersteunt. Deze basis staat bij elektrofysiologen bekend als de septale isthmus, en het is hier waar radiofrequente ablatie van het trage pad wordt uitgevoerd bij patiënten met AV nodale reënterende tachycardie.14
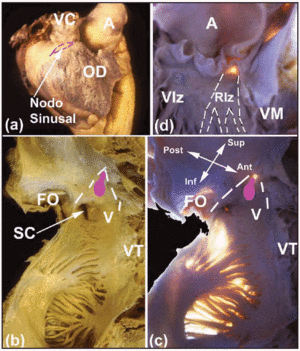
Fig. 2. A: lateraal epicardiaal aanzicht van de rechterboezem met de plaats van de SA-knoop aangegeven door de stippellijn in roze. B en C: endocardiale aanzichten (normaal en met transilluminatie) van de achterste en septale wanden van de rechterboezem om de ovale fossa (OF) en de grenzen van de driehoek van Koch (onderbroken witte lijnen), de pees van Todazo (TT) en de insertie van de septale cusp van de tricuspidalisklep (TV) te tonen. De vestibule (V) van de rechterboezem en de orifice van de sinus coronaris (CS) vormen de ondergrens. De plaats van de AV-knoop wordt aangegeven door een ovale knobbel (roze). D: Linkerventrikelaanzicht om het membraneuze septum te tonen (transilluminatie). Dit is het punt van ontstaan van de bundel van His en zijn continuïteit met de rechter en linker bundeltak. De linker bundeltak (LBB) is gemarkeerd met witte stippellijnen. A staat voor aorta; AA voor atriale appendage; VC voor superieure vena cava; RV voor rechter ventrikel; LV voor linker ventrikel; MV voor mitralisklep.
De voortzetting van de AV-geleiding gebeurt via de doordringende bundel van His, het enige deel van de geleidingsas dat de CFB doorboort. De CFB wordt gevormd door de vereniging van het bindweefsel van de aorta- en mitralisklepbladen met het septale blad van de tricuspidalisklep – de zogenaamde rechter fibreuze trigon – en het membraneuze deel van het interventriculaire septum. In veel harten van zoogdieren is de trigon vezelig, maar runderharten hebben een centrale massa bot of kraakbeen (het os cordis). Het vezelige weefsel van het hart van de potvis is daarentegen zeer los. Het membraneuze gedeelte of septum, dat in lengte kan variëren, is een goede gids voor het lokaliseren van de AV-bundel van Zijn. Deze verschijnt boven dit membraneuze gedeelte na het kruisen van de juiste vezelige trigon (figuur 2d), en verdeelt zich vervolgens in de linker en rechter bundel takken. De rechtertak gaat door de septale musculatuur aan de basis van de mediale papillaire spier van het rechter ventrikel. Hij wordt dan een dunne streng die diep doordringt in de septomarginale trabeculatie of moderatorband die de mediale en anterieure papillaire spieren verbindt. De oorsprong van de linkertak ligt onder de commissuur tussen de rechter en de niet-coronaire cuspen van de aortaklep; hij daalt vervolgens af door het subendocardium van het interventriculaire septum (figuur 2d). Het traject is soms zichtbaar door de glanzende vezelige lamina die het omhult. Het proximale deel van de linkertak is veel langer dan dat van de rechtertak. Af en toe wordt bij foetale of zuigelingenharten een derde tak gezien, die “doodlopende weg “15 wordt genoemd en die de bundel van His in anterio-superior richting voortzet naar de wortel van de aorta.
Structuur van de knopen en het atrioventriculaire geleidingssysteem
Studies waarbij de gebruikte histologische technieken vergelijkbaar waren met die van Tawara2 en latere werkers zoals Davies16 en Truex et al17 (om er maar een paar te noemen) hebben aangetoond dat het CS van de mens is gerangschikt op een manier die vrij veel lijkt op die van andere zoogdieren (met lichte variaties tussen soorten en tussen harten). Tawara2 rapporteerde de scheiding van de gespecialiseerde myocyten van de normale of werkende myocyten door een dun vel bindweefsel dat zichtbaar is onder de lichtmicroscoop, en hierop zijn de criteria gebaseerd die Aschoff18 en Mönckeberg19 hebben voorgesteld voor histologische identificatie van het gespecialiseerde myocardium. Eenvoudig gezegd onderscheiden gespecialiseerde myocyten zich van werkende myocyten wanneer zij onder de lichtmicroscoop worden bekeken, en kunnen zij van de ene histologische sectie naar de volgende worden “gevolgd”. In zijn monografie gaf Robb8 er de voorkeur aan om het geleidende weefsel te definiëren met de term “verbindend” in plaats van “geleidend” systeem, omdat histologische preparaten de celmorfologie beter definiëren dan de functie. Hij constateerde ook verschillen in de textuur van het gespecialiseerde myocard afhankelijk van de versheid van het autopsiemateriaal en de gebruikte fixatie- en kleuringsmethoden. Tawara2 was zich hiervan bewust en wees op de heterogeniteit van de gespecialiseerde myocytenmorfologie, zelfs in histologische coupes van het hart. Binnen een bepaalde soort houden de duidelijkste verschillen verband met de leeftijd van de onderzochte individuen.20 De laatste jaren zijn verschillende moleculaire en immunohistochemische merkers gebruikt om het geleidingsweefsel in embryonale harten van mensen en andere zoogdieren te lokaliseren. Er is echter geen specifieke marker gevonden die dit weefsel bij de volwassen mens kan markeren.
In het normale menselijke hart voldoen de SA- en AV-knopen niet aan de criteria van Aschoff en Mönckeberg18,19 omdat zij niet elektrisch van het omringende myocard geïsoleerd zijn door bindweefsel of vetweefsel. Integendeel, ze in contact komen met atriale werkende vezels na een klein gebied dat bestaat uit overgangscellen. In de SA-knoop maakten Keith en Flack5 onderscheid tussen de sinus- en de werkcellen. Tawara2 wees echter op de moeilijkheden die hij ondervond bij het onderscheiden van de cellen van de AV-knoop van die van de bundel van His. Hij stelde daarom voor dat het verschil tussen beide zuiver anatomisch was. Op basis van deze definitie wordt het gedeelte van de CS dat volledig omhuld wordt door de CFB de penetrerende bundel of bundel van His genoemd (figuur 3a). Het atriale gedeelte vanaf het proximale geleidingssysteem tot aan de bundel van His wordt de AV-knoop genoemd (figuur 3b). Dit anatomische onderscheid is logisch omdat de isolatie van de doordringende bundel van His voorkomt dat deze rechtstreeks contact maakt met de elektrische activiteit van het afferente atrium. Eventuele atriale activiteit moet dus vooraf via de AV-knoop worden geleid.
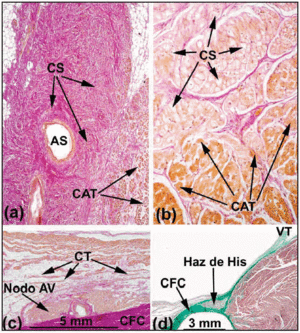
Fig. 3. Sagittale histologische doorsneden van de sinoatriale knoop (SA) van het menselijke (a;x10) en varkenshart (b;x40) gekleurd met de van Gieson methode. Let op het contact tussen sinuscellen (SC) en werkende atriale cellen (WAC). De sinuscellen zijn duidelijker en ingebed in een grotere hoeveelheid bindweefsel (rood). c: Van Gieson-gekleurde doorsnede van het middengebied van de driehoek van Koch. Let op de vorm van de compacte AV-knoop en de overgangscellen (TC) in contact met het convexe oppervlak van de compacte knoop. d: Masson’s trichroom-gekleurde sectie die de doordringende bundel van His toont, omgeven door fibreus weefsel (groen) van de CFB. SNA staat voor sinusknoopslagader; CFB voor centraal fibreus lichaam; TV voor tricuspidalisklep.
De intrinsieke functie van de SA-knoop is de bron te zijn van de hartimpuls. De SA-knoop bij de mens is een gewelfde of fusiforme structuur. Histologisch is hij samengesteld uit cellen die iets kleiner zijn dan normale werkcellen en die in bundels zijn gerangschikt. Deze lopen door elkaar zonder ruimtelijke orde, kleuren zwak, en zijn ingebed in een dichte bindweefselmatrix (Figuren 3 a en b). Met de leeftijd neemt de hoeveelheid bindweefsel toe ten opzichte van het gebied dat door de knoopcellen wordt ingenomen.21 Aan de periferie van de knoop zijn gespecialiseerde cellen vermengd met die van het werkend myocard (Figuren 3a en b). Bovendien zijn er meerdere radiaties of extensies beschreven die interdigiteren met het werkende atriale myocard. Deze dringen intramyocardiaal door in de terminale kam, en de vena cavae superior en inferior. De SA-knoop is gegroepeerd rond een slagader die bekend staat als de sinusknoopslagader, die centraal of excentrisch in de knoop kan lopen. In 29% van de menselijke harten vertakt deze slagader zich binnenin de knoop.11 De SA-knoop is ook nauw verbonden met het autonome zenuwstelsel. Er is gesuggereerd dat de meerderheid van deze zenuwvezels parasympatisch zijn, terwijl de sympatische vezels geconcentreerd zijn rond de bloedvaten van de knoop.23
De inherente functie van de AV-knoop is het vertragen van de hartimpuls. Bij de mens heeft deze knoop een compact gedeelte en een gebied met overgangscellen. Het eerste is halfovaal en ligt over de CFB (figuur 3c). In de doorsneden dicht bij de basis van de driehoek van Koch, verdeelt het compacte deel van de knoop zich in twee verlengstukken of verlengstukken. De slagader die de AV-knoop doorbloedt, bevindt zich gewoonlijk tussen deze twee. De lengte van deze verlengstukken varieert van hart tot hart.24 De grootte van de overgangscellen ligt tussen die van de AV-knoop en de atriale werkcellen in. Zij zijn omgeven door een grotere hoeveelheid bindweefselcellen dan die welke de werkcellen bedekken, maar zij zijn niet geïsoleerd van het aangrenzende myocardium. Zij vormen veeleer een soort brug tussen het werkend myocard en het knooppuntmyocard, en verzamelen elektrische informatie van de atriale wanden en geven die door aan de AV-knoop.
Er bestaat onenigheid over de wijze waarop de impuls van de SA-knoop de AV-knoop bereikt. Sommige auteurs hebben het bestaan gesuggereerd van gespecialiseerde traktaten tussen hen.25 Onze studies ondersteunen dit idee niet, maar zijn voorstander van de hypothese dat de werkende spiervezels zelf (en hun geometrische rangschikking in de atriale wanden) verantwoordelijk zijn voor geleiding die sneller is in sommige gebieden van het atrium dan in andere.26
De AV-knoop gaat distaal verder met de doordringende bundel van His (figuur 3d), hoewel er kleine verschillen zijn in termen van cellulaire rangschikking tussen deze twee structuren, met inbegrip van de rangschikking van de bundel van His cellen op een meer parallelle manier. De verklaring hiervoor zou morfologisch kunnen zijn: de bundel His begint omgeven te worden door het bindweefsel van de CFB, en wordt zo een geleidend kanaal dat informatie naar de ventrikels brengt.
De AV-knoop van de hond is kleiner dan die van de mens, maar heeft een langere doordringende bundel His.27 Sommige auteurs28 interpreteren dit zo dat een deel van de AV-knoop van de hond binnen de CFB ligt. Bij het konijn beschrijven andere auteurs29 een deel van de bundel His alsof het een deel van de AV knoop is, maar dit is een vergissing (Figuren 4a-d). Het meest opvallende morfologische verschil tussen de AV knoop van de hond en die van het konijn en de mens is dat de eerstgenoemde niet door overgangscellen wordt bedekt. Bij ratten (met een hartslag in rust die 10 maal sneller is dan bij de hond of de mens), is de AV knoop verhoudingsgewijs vergelijkbaar met die van de hond, maar de CFB is kleiner.
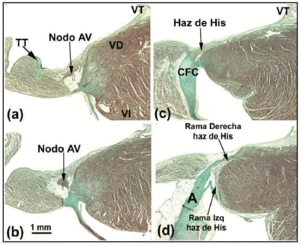
Fig. 4. Deze samengestelde figuur toont de atrioventriculaire (AV) knoop plus de bundel van His en zijn rechter en linker bundeltakken bij het konijn. De horizontale balk in b vertegenwoordigt 1 mm (hetzelfde voor alle beelden). Masson’s trichroom kleuring. A geeft aorta aan; TT, pees van Todaro; RV, rechter ventrikel; LV, linker ventrikel; TV, tricuspidalisklep.
Wanneer het histologische traject van het geleidingssysteem wordt gevolgd in de richting van de doordringende bundel van His, is te zien dat deze in veel menselijke harten naar links draait, en op de gespierde kam van het interventriculaire septum tevoorschijn komt. Omgeven door bindweefsel van de CFB, kan de lengte van de bundel van His variëren alvorens zich te splitsen in de linker en rechter bundeltak. De eerstgenoemde tak cascadeert over de linkerzijde van het interventriculaire septum (Figuren 5a en c). De splitsing van de bundel van His lijkt op een jockey die hurkt boven de gespierde kam van het interventriculaire septum (Figuur 5a). Soms echter wijkt hij af naar links (Figuur 5c). Wanneer dit gebeurt, komt de rechtertak binnen in de binnenzijde van de musculatuur van het septum (Figuur 5b), en verschijnt in de rechterkamer in associatie met de insertie van de mediale papillaire spier.
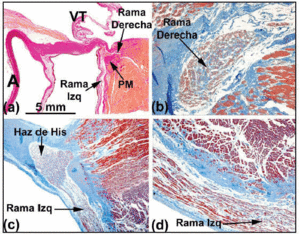
Fig. 5. a: van Gieson-gekleurde doorsnede die de bundel van His toont (menselijk hart) over het membraneuze gedeelte (MP) van het interventriculaire septum. b: bij de oorsprong is de rechtertak in dit hart intramyocardiaal en omgeven door bindweefsel (blauw) (Jones’ trichroomkleuring, x20). c: de splitsing van de bundel van His in dit hart is verplaatst over de linkerzijde van de spierkam van het interventriculaire septum, en daalt in de lengterichting af onder het endocard van de linker hartkamer (Jones’ trichroom, x5). d: let op de bedekkende bindweefselschede (blauw) van de linkertak (Jones’ trichroom, x10). A geeft aorta; E, endocardium; TV, tricuspidalisklep.
Langs hun proximale cursussen, de rechter en linker bundel takken zijn bedekt met een vezelige lamina (figuren 5b en d). Zoals Tawara2 liet zien (figuur 6a), is bij de mens de linkertak typisch verdeeld in drie fascikels met uitgebreide onderlinge verbinding. Deze fascikels worden vertakt in de ventriculaire apex, en uit te breiden tot het interieur van de twee papillaire spieren van de mitralisklep, maar ook terug langs de ventriculaire wanden in de richting van de hartbasis. Meer distaal, in de apex van de ventrikels van het menselijk hart, wordt het bijna onmogelijk om de vertakkingen van de Purkinje-vezels te traceren, omdat deze hun vezelige mantel verliezen en veel lijken op het werkende myocardium.
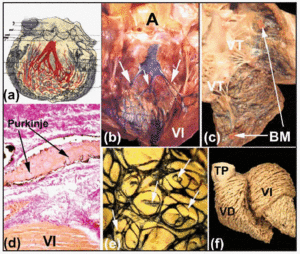
Fig. 6. a: diagram van Tawara dat de trifasciculaire rangschikking van de linkerbundeltak bij de mens toont. b en c: een vers kalfshart waarin de rechter en linker ventrikels zijn geopend. Subendocardiale injecties met Indische inkt onthullen de rechter en linker bundeltakken en het Purkinje netwerk. Let op in B de drie fascikels van de linker bundeltak (pijlen), en in C de moderatorband (MB). d: doorsnede van een kalfshart (van Gieson-kleuring, x100), verkregen na inspuiting van Indische inkt in het Purkinje-netwerk), dat aan de oorsprong omhuld is door bindweefsel (rood). e: subendocardiale rangschikking van het Purkinje-netwerk in de linker hartkamer van een kalf. Let op de elliptische opstelling van het netwerk en de uitlopers van de randen die het myocard doordringen (pijlen). f: dissectie van de ventriculaire spiervezels van een volwassen menselijk hart. Let op het verschil in rangschikking tussen de mediale en diepe lagen van de linker hartkamer. A geeft aorta; PT, pulmonale stam; LV, linker ventrikel; TV, tricuspidalisklep.
Subendotheliale injectie van Oost-Indische inkt is een van de methoden die worden gebruikt om deze vezelige vellen te observeren en om de subendocardiale verloop van de rechter en linker bundeltakken en hun vertakkingen aan te tonen in de harten van hoefdieren (figuren 6b en d). Onze studies van de harten van schapen en kalveren tonen aan dat deze enigszins verschillen van de menselijke harten. Kalfsharten zijn meer vergelijkbaar met menselijke harten in die zin dat de fascikels van de linker bundeltak meestal drie in aantal zijn en hun oorsprong vinden in het bovenste deel van het interventriculaire septum (figuur 6b). Schapenharten vertonen echter slechts twee fascikels, en deze verschijnen halverwege de lengte van de septale wand. In zowel schapen- als kalfsharten doorkruisen kleine spiertrabeculae de ventriculaire holte – de zogenaamde “valse pees” – die binnenin de distale vertakkingen van de His-takken naar de papillairspieren en de aangrenzende ventriculaire wanden voeren. Aan de rechterzijde van het hart is de moderatorband van zowel het schapen- als het kalverenhart slanker dan die van de mens, maar binnenin bevat hij altijd een uitloper van de rechterbundeltak (figuur 6c).
In de harten van hoefdieren is het subendocardiale Purkinje-netwerk ellipsvormig gerangschikt, zowel in de linker- als in de rechterkamer (figuur 6e). Bovendien ontstaan uit de omtrek ervan takken die de ventrikelwanden binnendringen en leiden tot nieuwe takken of anastamosen met andere takken (figuur 6e). Echter, intramurale takken van het Purkinje-netwerk zijn niet aangetoond in het menselijk hart.30
Een controversieel punt met betrekking tot het Purkinje-netwerk is het bestaan van overgangscellen tussen het werkende ventriculaire myocard en de Purkinje-vezels.31 De anatomische en immunohistochemische studies van Oosthoek et al,30 tonen aan dat, in runderharten, er een zeer kleine zone van overgangscellen is waar de Purkinje-vezels hun bindweefselbedekking verliezen. Dergelijke cellen zijn echter niet waargenomen in het schapenhart.32 Wanneer de Purkinjevezels hun bindweefselbedekking verliezen, gaan er elektrische impulsen van het CS naar de werkende myocyten van de ventrikels. De ruimtelijke oriëntatie van de werkende myofibrillen in de ventrikelwanden bepaalt de anisotrope aard van ventriculaire geleiding (figuur 6f).
CONCLUSIES
Hoewel er verschillen bestaan tussen de soorten, de structuur van de knooppunten, evenals die van de rest van het menselijke AV geleidingssysteem, is vergelijkbaar met die van veelgebruikte proefdieren. De SA-knoop, de structuur die de hartimpuls genereert, bevindt zich aan één uiteinde van de rechterboezem. Impulsen van deze knoop reizen posterieur in de atriumwanden door een ingewikkelde maar precieze ruimtelijke rangschikking van werkende atriumvezels totdat zij het eind van het atrium bereiken. Aan dit einde ontvangen de overgangscellen van de AV-knoop de impulsen en vertragen deze voordat zij via de bundel van His worden doorgegeven. Deze laatste doorkruist het isolerende vezelvlak tussen boezems en kamers en geeft de impuls via twee takken (de rechter en linker bundeltak) door aan de corresponderende kamers. Elk van deze takken is geïsoleerd door een verbindingsmantel van werkende ventriculaire myocyten. Door deze opstelling is contact tussen de gespecialiseerde en werkende myocyten alleen mogelijk aan de distale vertakkingen van de bundel van His. Op deze wijze is het AV-geleidingssysteem, dat bijna 100 jaar geleden door Tawara2 grotendeels werd beschreven, gestructureerd om orde te brengen in de overdracht van cardiale impulsen. Kennis van de structuur en de plaats van specifiek geleidingsweefsel in het hart zou kunnen helpen oplossingen te vinden voor verschillende stoornissen in het hartritme.
Leave a Reply