South Americaブッシュドッグ Speothos venaticus

bush dog – © Gerald L Zuercher
Amazonian Canids Working Group – Karen DeMatteoとFernanda MichalskiはAmazonian Canids Working Groupのコーディネーターである。 このワーキンググループは、コビトイヌ、カニクイキツネ、ブッシュドッグ、そして南米のキツネの4種に焦点を当てています。 アマゾンのイヌ科動物は世界中の多くの肉食動物と同様、生息地の喪失、水力発電ダムの拡張、獲物の違法狩猟、家庭犬による病気など、さまざまな直接・間接的脅威によって長期的な生存が脅かされているのです。
プロジェクト
- Proyecto Zorro Pitoco: アルゼンチン・ミシオネス州のブッシュドッグと共存する生物多様性について、非侵襲的な手法で新たな知見を得る
- Camera trap wildlife monitoring in and around Madidi National Park, ボリビア
- ミシオネラ森林の野生動物のビオトープにおける新熱帯蠕虫症およびその他の寄生虫の調査、人獣共通感染症の潜在的病巣として
- アマゾンにおける肉食動物に対する気候変動の影響
関連リンク
- Flickr: 3616>
- 2011 IUCN Red List Assessment – Bush Dog
Reports / Papers
- Residents of the forest – Canids from Amazon
- 2004 Status Survey & Convervation Action Plan – (アマゾンの野生イヌ科動物について) 南米
- なぜブッシュドッグは他の犬とそんなに違うのか
- カメラに収められた珍しいアマゾンのブッシュドッグ
- 救助犬は今、他の(もっと野生の)犬を救うのを手伝っている
- 驚くべき種。 bush dog
その他の名称
English: ブッシュドッグ、サバンナドッグ、ビネガードッグ
フランス語:Bush Dog、Vinegar Dog。 Chien Des Buissons, Zorro
スペイン語; Castilian:Cachorro Vinagre, Guanfando, Pero Selvático, Perrito Venadero, Perrito de Monte , Perro De Monte, Perro Grullero , Perro Vinagre , Perro de Agua, Perro de la Selva, Umba, Zorrito Vinagre, Zorro Pitoco , Zorro Vinagre
German.Odyssey:German’s Dogs, Zorro Pinagre, Zorrot, Zorrot Vinagre, Zorrot Vinagre, Zorrot Vinagre, Zorrot Vinagre, Zorrot Vinagre, Zorrot Vinagre, Zorrot Vinagre, Zorrot Vinagre Waldhund
イタリア語:Itticione、Speoto
ポルトガル語:Portuguese: Cachorrinho, Cachorro-do-mata, Cachorro-do-mato-cotó, Cachorro-do-mato-vinagre, Cachorro-pitoco
Justification
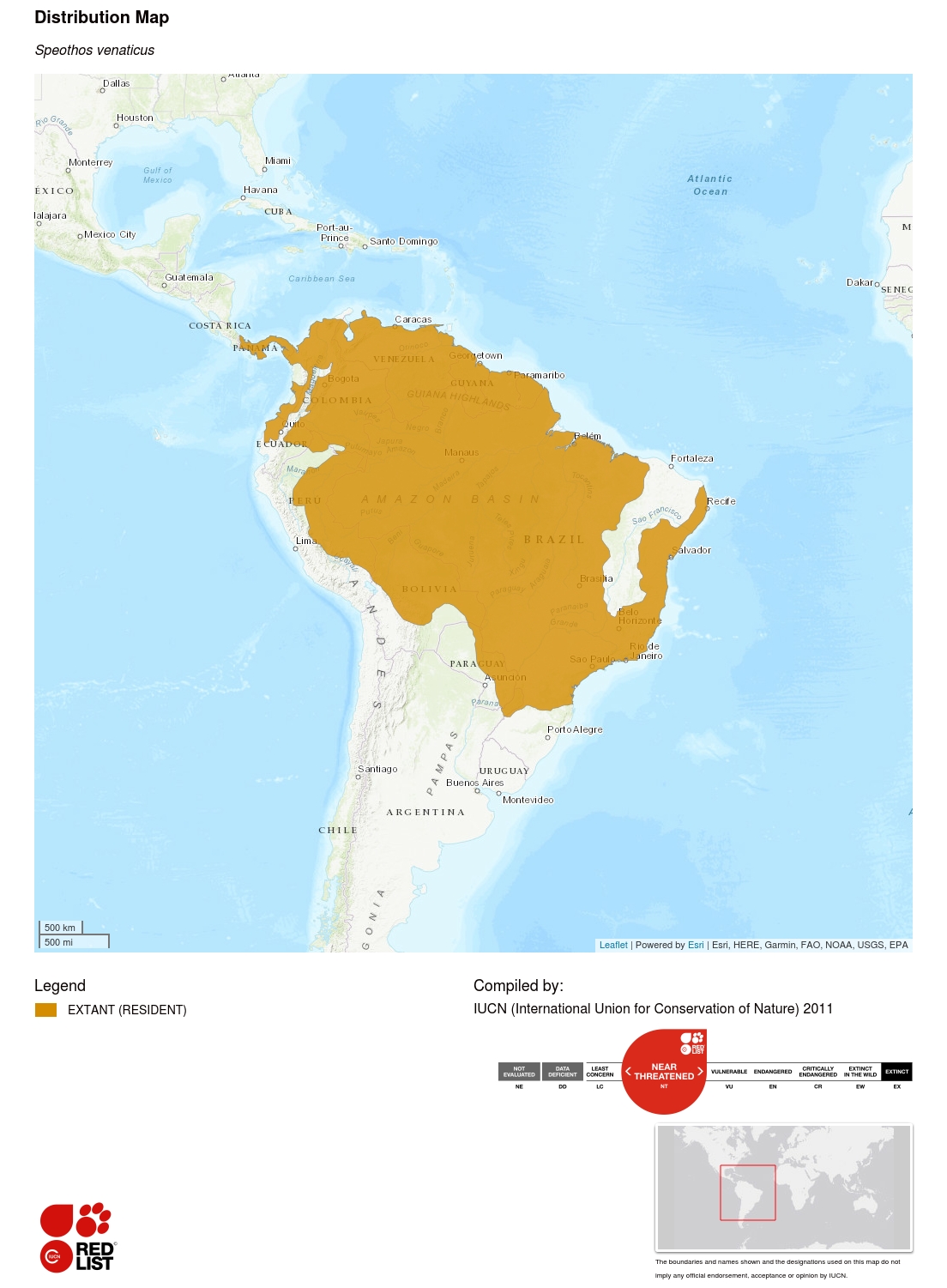
ブッシュドッグは広範囲にわたっているが、そのすべての範囲でまれと思われている。 この種は野生で見つけるのが非常に困難であることが証明されており、個体数の傾向を推定するのが困難である。 断片化された、あるいは変更された生息地での発生が確認されている広範囲に分布する種は、一般的に安定した個体数を持つと予想されるが、現地からの予備的な証拠は、これらの地域がブッシュドッグの生態学的要件を変更し、結果として集団を支えるために必要な最小面積が増加する(そしてそれによって、種が他の脅威に直接または間接的にさらされるリスクが高まる)可能性を示唆するものだ。 この種は過去12年間(推定世代数=4年)で20~25%減少したと推定され、基準A2のもとで弱者として登録される閾値に近いことから、ここでは近絶滅危惧にリストアップされている。 この減少は、進行中の生息地の損失と断片化、違法な狩猟と家庭犬による捕食による餌の豊富さの減少、およびイヌ科の病気に起因するリスクの増加など、複数の脅威に起因している。 また、アンデス山脈の西にあるエクアドルとコロンビアには、現在も孤立した亜集団が生息している可能性がある。 歴史的な分布はコスタリカまで北上していた可能性があり(Rosa and Nocke 2002)、そこではこの種がまだ生存している可能性がある。 現在の分布図は、肉食獣の生物学者および文献による種の存在に関する広範な調査結果(n = 399の歴史的場所)と、この種に適した領域を予測する生物気候モデル(DeMatteo and Loiselle 2008)を用いて作成されたものである。 後者の著者らは、ブッシュドッグを生息させるのに適したと予測される総面積は14,445,000 km²のオーダーであり、このうちの77%が現在の範囲内で発生すると推定している。
個体数の傾向:減少中
生息情報
広い分布範囲と様々な生息地で見られるにもかかわらず、ブッシュドッグはその範囲全体で自然には珍しい存在と思われています。 DeMatteo(2008)による最近の調査では、ブッシュドッグの分布にある大多数の国が、この種の状態を希少または不明と報告している(希少:エクアドル、希少または不明:エクアドル。 アルゼンチン、ボリビア、ブラジル、フランス領ギアナ、パラグアイ、不明。 パナマ、ベネズエラ)、2カ国だけが普通種と報告している(ガイアナ、ペルー)。 この種はカメラトラップではめったに記録されないが、検出されないことを解釈する際には注意が必要で、これは実際にこの種がこの手法に関連する特徴を積極的に避けていることに関連するかもしれない(例.
ブッシュドッグの人口統計学的データはその範囲全体で不足しており、個体数の推定はいくつかの地域でのみ報告されている。 <Misiones Argentinaで100頭 (DeMatteo 2008), >Bolivia (DeMatteo 2008), >Cusco PeruのCamisea River地域内の4,022 km² or one individual/4 km² で1000頭 (DeMatteo 2008), 0.04 個体/km2 (B. Beisiegel pers. comm) in Brazil, Mato Grosso Brazilで一部断片化しているセラードでは 0.025 個体/ km2 (E. B. Deimatto 2008), Bolivia で 1,000 頭 (D. Deimatto 2008), >Bolivia では 1,000 頭を超える個体が生息している (Deamatto 2008).S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima, and S. Klorfine, pers. obs.), ブラジル、パンタナールで 15.8 個体 /394 km² または 0.04 個体 /km² (Lima, Jorge et al. 2009) そしてブラジル南部アマゾンでの 0.001 個体 /km² (Michalski 2010)であっ た。 生息適地が約11,000,000km²(DeMatteo and Loiselle 2008)、平均個体数が約0.01個/km²とすると、推定生息数は110,000個で、その約半数が成熟していると考えられるが、断片化の影響、保護の度合い、広い分布における変動からこれは過剰推定であることが示唆される。
ブッシュドッグの分布全体では、2つの国(ガイアナとパナマ)で不明、2つ(パラグアイとベネズエラ)で減少、1つ(フランス領ギアナ)で不明または安定、2つ(エクアドルとペルー、特にクスコペルー)で安定と報告されている(デマッテオ2008年)。 全範囲にわたって、生息地の断片化(都市化と農業)のレベルの増加、違法な密猟による獲物集団への脅威、致命的な可能性のあるカンジダ関連疾患にさらされていることから、安定または増加傾向にある可能性は極めて低いと思われる。
生息地と生態情報
ブッシュドッグは先住民によって、一般的に水源、特に餌の密度が高くなりそうな小さな川の近くで発生するという文脈で、生息地の一般主義者であると報告されている(Zuercher et al. 2005; E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima, and S. Klorfine pers. obs.; M. Swarner, pers. comm)。 ブッシュドッグは低地(1,500 m以下)の原生林や回廊林(Defler 1986)、半落葉樹林、季節的洪水林などの森林生息地で観察されている(Aquino and Puertas 1997)。 また、ブラジルのセラード生息地(Silveira他 1998; Oliveira 2009, C. Brady pers. 最近の報告書では、ブッシュドッグを3つの独特な生息地:カーティン ガ、チャコ、海岸沿いのマングローブで観察していることが記載されている(DeMatteo and Loiselle 2008)。 場合によっては、森林の生息地から数km離れた場所で観察されることもある(Silveira et al.1998)。 また、本種は二次林、牧場地(M. Swarnerの私信)、断片化したセラード牧場地(E. S. Lima, K. E. DeMatteo, R. S. P. Jorge, M. L. S. P. Jorge, J. Dalponte, H. S. Lima, and S. Klorfine pers. Jácomo pers.), disturbed areas (DeMatteo and Loiselle 2008, Oliveira 2009), and fragmented forest areas (Michalski and Peres 2005, Michalski 2010).
ブラジルのPantanal (Lima, Jorge et al.) でも生息地選択の優先は見いだせず、また、Pantanalでは、PantanalとPantanalに生息し、Pantanalでは、L. 2009)、パラグアイの上部パラナ大西洋岸森林-セラード混合地帯(Zuercher et al.2005)のいずれでも生息地選択の嗜好性は認められなかった。しかし、一部断片化したセラードでブッシュドッグのグループが改変地(農地や放牧地)に対して手つかずのサバナや森林生息地を好むことを示す証拠がいくつか見つかり、地域や入手可能性によって生息地の利用や嗜好性が異なる可能性を示している(E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima, and S. Klorfine pers.obs.). Michalski(2010)はアマゾン南部の断片化された地域にブッシュドッグが存在しないと報告しているが、他の研究はブッシュドッグが存在する可能性が高いという強い証拠を示している(DeMatteo and Loiselle 2008; Oliveira 2009; E.S.Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima, S. Klorfine pers. obs)、これらの地域での使用の強度と生態的要件への影響は不明である。 現地で得られた予備的データによると、生息地の断片化の程度が増すと、ブッシュドッグが必要とする面積も増えることが示唆されている(E.S. Lima私信)。このデータはブラジルのセラード生態系でのみ得られているが、すべての生息地タイプで当てはまる可能性があり、餌の密度と生息地の断片化の間の負の関係の反映であることが推測される。 ブッシュドッグの歴史的分布(n > 250地点)の再分析では、約4分の1(20%)が断片化/改変された生息地と関連しており、推定値を単一個体を支えるために必要な面積から繁殖集団の面積に拡大すると、改変された生息地の割合が増加した(DeMatteo and Loiselle 2008)。
報告されている生息域の広さの推定値のばらつきは、生息地の完全性と餌の密度に関連していると考えられる:150 km² (Upper Paraná Atlantic Forest; Beisiegel 1999), ca. 16 km² (Beisiegel and Ades 2004), 140 km² (Fixed Kernel 95%) セラードのブッシュドッグの1グループ (n = 2-4 個体) (E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S.. J., K.E. Jorge, K.E. DeMatteo) Lima, and S. Klorfine, pers. obs.).
予備的なフィールドデータは、ブッシュドッグが1つ以上の仔のいる家族グループで存在するという飼育下研究を支持している(DeMatteo 2008, Michalski 2010, E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima, and S. Klorfine, pers. obs.); しかしながら、単独で行動する個体が様々な生息環境の中で定期的に観察される。 野生での繁殖は季節に関係なく行われる (DeMatteo 2008)。 本種は多くの肉食獣のような真のテリトリーではなく、半遊動的な移動パターンを持つようである(DeMatteo 2008, E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima, and S. Klorfine, pers. obs.) またペルーとブラジルではAtelocynus microtisと同所的に生息することが記録されている (Leite Pitman, Beck et al. 2003, Michalski 2010)。 推定世代数は4年[(0.5×生殖期間6年)+1年で性成熟(飼育下;Porton他1987)]。
予備調査データは、食性に関する逸話的報告やブッシュドッグが非常に肉食であるという考え(Deutsch 1984, Peres 1991)を支持しているが、地域や生息地の種類による餌の好みがあり、季節変動も重なったと思われる。 ブラジルのパンタナル地区では、主食は九帯アルマジロ(Dasypus novemcinctus、94%)で、小型哺乳類、アグーチ(Dasyprocta azarae)、鳥は少量であったが、果物は記録されていない(n = 17、匂いと痕跡で特定;Lima、Jorge et al.2009). パラグアイの大西洋岸森林内では、食物の大部分はアグーチとパカ(Cuniculus paca)で構成され、小哺乳類、げっ歯類、爬虫類、無脊椎動物およびセクロピアの果実は少量であった(n = 11、ミトコンドリアDNAにより同定;Zuercher et al.2005)。 肉と果物(バナナとパパイヤ)が入手できるブラジル南東部の繁殖センターでは、動物は果物と肉を同じ割合で食べている(L. Saboia, 私信)。 家禽の捕食は時々起こると報告されている(DeMatteo 2008)。
脅威情報
以下のようないくつかの深刻な脅威が認識されている。 1) 大規模農業(例:大豆)、牧草地への土地の転換、および単一栽培樹木の大規模プランテーション(例:…)による人間の侵入と無傷の生息地の損失。 2)違法な密猟や家庭犬の捕食による餌の豊富さの減少、3)家庭犬から致死性の病気に感染するリスクの増加(人間集団や狩猟犬に近接している(DeMatteo 2008)。 しかし、フィールドの証拠は、病原体が家庭犬によって伝染する可能性があることを支持し、その影響は、主にこの種の集団生活のために、潜在的に壊滅的であることができる(Mannら、1980年、Steinelら、2008年)。 2001, Leite Pitman, Nieto et al. 2003, Jorge, Morato et al. 2007, Jorge, Nunes, et al. 2007, DeMatteo 2008, Jorge et al. 2008, E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J.Dalponte, H.S. Lima, and S. Klorfine, pers. obs.). E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima, S. Klorfine (pers. obs.) は、疥癬と思われる全身脱毛が徐々に野生ブッシュドッグのグループに広がり、ついにはすべての個体を死亡させたことからこの効果を確認したという。 疥癬に加え、パルボウイルスと狂犬病も野生個体群に悪影響を及ぼす病気として報告されている(Mann et al.1980、DeMatteo 2008)。 本種はDioctophyma renaleおよびAmphimerus interruptus(博物館所蔵 – Vierira et al.2008)、Lagochilascaris sp.(VolcánおよびMedrano 1991)に感受性があり、Echinococcus vogeli(Cestoda: Taeniidae) の宿主として確認されている (Rausch and Bernstein 1972). 飼育動物は、パルボウイルス(Janssen et al. 1982)、ワクチン誘発性犬ジステンパー(McInnes et al. 1992)、リーシュマニア(Lima, Fattori et al. 2009)、スピロカ ルピ(Rinas et al. 後者では、大家族の15匹のうち13匹が、誤って与えた感染したコアティからカンピロバクターに感染し、重度の出血性腸炎で急死した(L. Saboia私信)。 このように短期間に複数の動物が死亡したことは、様々な病気に対する種の感受性と集団生活種が病原体に対して敏感であることを強調するものである。 人間居住区への接近や狩猟犬による保護区への侵入が増加しているため、病気にかかるリスクは高い可能性がある。 また、人間から独立して定期的に獲物を狩る、ワクチン接種をしていない野良犬や半野良犬の飼い犬が病気にさらされる可能性もある(K. DeMatteo pers.) DeMatteo (2008)は、ブラジルでは家畜よりもブッシュドッグと飼い犬の間で問題のある相互作用が起きていると指摘しており、この種に対する病気の脅威が強調されているにすぎない。
Use and Trade Information
ブッシュドッグは5カ国(アルゼンチン、ブラジル、パナマ、ペルー、ベネズエラ)で撃たれ、2カ国(パナマとペルー)で罠にかかったと報告されている(DeMatteo 2008, F. Michalski unubl. data)。 ペルーでは、これらの出来事は、先住民がブッシュドッグを罠にかけたら食べるか、伝統的な狩猟で集める稀な機会があれば食べると報告している。しかし、ベネズエラでは、銃殺はその希少性から、時折トロフィーの収集と関連していると報告している (DeMatteo 2008).
保全活動情報
この種はその範囲内のいくつかの保護区で見られるが、パッチ状の分布で非常に低い密度で生息している。 コロンビア(法律番号848:1973)、エクアドル(法律番号74:1981)、フランス領ギアナ(法律番号JO19860625:1986)、パナマ(法律番号2-80:1980)、パラグアイ(法律番号18796:1975)およびペルー(法律番号5056:1970)で狩猟が禁止されています。 アルゼンチン(法律番号22.421:1981)、ボリビア(法律番号12301:1975)、ブラジル(法律番号5197:191967)、ベネズエラ(法律番号276:1970)では狩猟と取引が規制されている。 ガイアナとスリナムの情報はない。 残念ながら、その範囲の多くの部分で、指定された保護区域を管理し、既存の野生生物法を施行するためのリソースが不十分である。
ワシントン条約の付属書Iに含まれる。
ブッシュドッグは飼育下で発生し、アジア(日本)、ヨーロッパ、ブラジル、北アメリカを含む国際繁殖プログラムの一部で成功している(Buck 2009)。 再導入の試みは知られていない。
ブッシュドッグの個体数の推定と人口統計学的データは、その範囲全体ではまだほとんど知られていない。 これは個々のグループの社会力学、特に若者の分散と他のグループとの相対的な使用領域(重複または別々のホームレンジ)の理解にも及ぶものである。 この種はかつて森林に依存していると考えられていたが、現在では開放的で断片化した生息地で観察されることが多くなっている。 食性に関する予備的データは主に肉食性であることを裏付けているが、食性に関する季節的変化や地理的変動は評価される必要がある。 病気の影響、特に家庭犬からの感染動態を野生の個体群でどのように管理し、最小限に抑えることができるかを決定する必要がある。
カメラトラップやトランセクト調査などの標準的な調査技術を使用して種の存在の証拠を見つけることは難しいことが分かっている(Beisiegel 2009, DeMatteo et al.2009, Michalski 2010)。 特定の場所への誘引力を高める人工的な香りのルアーの使用は、野生のブッシュドッグでは成功しなかった(Zuercher他、1999年)。 しかし、カメラの設置高さの調整、捕獲努力の増加、ロングコールの発声再生と同種の尿の同時使用など、この種に対するこれらの手法の有効性を高めることができるいくつかの方法論的調整がある(DeMatteo et al.2004)。 部分的に分断されたセラードでの限定的な野外試験(2004-2005)では、プレイバック、尿、足掛けトラップは成功しなかった(K. DeMatteo未発表データ)。 しかし、種の密度、生息地の多様性(森林とセラード)、餌の密度がどのように技術の有効性を変化させるかについては、さらなる試験が必要である。
3つの非侵襲的技術(匂い探知犬、糞便DNAスクリーニング、GIS技術)の組み合わせを用いた予備的かつ進行中の研究は、この種に対して成功することが示されており(DeMatteo他、2009、未発表)、さらなる地域と生息地に拡大されるべきである。 この一連の技術により、特定の場所にこの種を呼び寄せる必要がなくなり、生息地の利用(無傷と断片化)、個体数の状況、最小利用面積、他の肉食獣とのニッチの重複/分離など多くの生態的変数について洞察することができる
。
Leave a Reply