Frontiers in Plant Science
The Different Types of Heterochrony
1870年代にエルンスト・ヘッケルは、進化にとって最も重要なメカニズムとして、祖先に対する子孫の発生における時間変化と空間変化を挙げた(ヘッケル、1875年)。 ヘッケルは空間的変化をヘテロトピー、時間的変化をヘテロクロニーと名付けた。
しかし、ヘテロクロニーの意味はヘッケルが最初にこの言葉を作って以来変化してきた。 ヘッケルは、彼のよく知られた「生物遺伝学的法則」(発生過程の順序(個体発生)は、種の進化史における順序(系統発生)をほぼ再現するという法則)からの逸脱を指して、ヘテロクロニーという用語を使った(ヘッケル、1875年)。 したがって、ヘテロクロニーとはもともと、ある生物の発生系列において、ある特徴が現れる時期が、その生物の系統発生における順序と比較して変化していることを意味する。 20世紀半ば、デビア(1951)はヘテロクロニーを再現性から切り離した。 彼は、近縁の2つの種を比較したときの発生時期の違いをヘテロクロニーと呼び、ヘテロクロニーがどのように生物間の多様性を生み出すかを説明したのである。 Gould(1977)は、ヘテロクロニーを「祖先にすでに存在する特性の相対的な出現時期や発達速度の変化」と定義し、ヘテロクロニーを検出するために、発達の時期ではなく、相対的な大きさや形状の変化を重視し、再捕捉の概念と再度関連づけた。 21世紀に入り、スミスは、最終的な大きさや形の変化を重視する「成長ヘテロクロニー」と、ヘッケルとデ・ベールの本来の用法に近く、発生イベントのタイミングの変化によって表現型の変異を説明できる「シーケンスヘテロクロニー」の2種類を定義することがより有用であると提案した(スミス、2002、2003;キーテとスミス、2014年)。
シーケンスヘテロクロニー(以下、ヘテロクロニー)は、小児型と変態型の2つに分類される。 祖先の発生と比較すると、幼形化は幼体または単純な結果をもたらし、一方、変態は成体またはより複雑な表現型をもたらす。 これらの2つのカテゴリーの異時性は、Alberchら(1979)が提案したように、発生プロセスの開始、オフセット、速度のタイミングのばらつきに起因する可能性がある。 この変動は6つの異なるタイプの異時性をもたらす(図1)。 幼形化は、発生過程の早期終了(前駆)、発生過程の開始の遅れ(後置)、発生速度の低下(新生)のいずれかに起因することがある。 変態は、発生の終了が遅い(hypermorphosis)、開始が早い(pre-displacement)、または発生速度が速い(accelerated)ため、発生期間が長くなった結果である(図1A)。 植物の胚発生や植物体形成における過形態と小形態の仮定的な例を図1B,Cに示す。
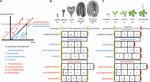
FIGURE 1. ヘテロクロニーの種類。 (A)6種類のヘテロクロニーの模式図。 黒線は祖先の個体発生において、ある発生段階に到達するのに必要な時間を表す。 青線は3種類の異形化:前駆(早発型)、後駆(遅発型)、新生(遅発型)。 赤線は変態の3つのタイプ、すなわち超変態(オフセットが遅れる)、前置換(発症が早い)、加速(発達速度が速い)を示している。 (A)の図面はAlberchら(1979)とGeuten and Coenen(2013)に基づいている。 (B) 胚発生におけるヘテロクロニーシナリオ。 ここでは、シロイヌナズナの胚発生を仮想的な祖先の発生とし、説明のために4つの段階に分けている。 胚が最初の2枚の葉と2枚の子葉を作る第5段階(胚実生と表記)は、周形成期の胚発生の最終段階として提案されており、一方、前形成期の胚発生は心臓段階で終了すると予想される。 (C) 植生発生のヘテロクロニーシナリオ。 この場合、シロイヌナズナの植物体形成は、開花前に茎に尾状葉・繁殖葉を作り続ける(上に描く)ことで、尾状葉を作らずに成体期に開花する仮想的な祖先と比較して、周形態的な植物体形成の仮想的なケースを表している。
これらの分類は発生時期の変化を説明するのに便利ですが、実際の例を考えると、異なる種類の異時性を区別するのは難しいことがよくあります。
植物の進化と多様化におけるヘテロクロニー
陸上植物(胚葉植物)は、約4億7000万年前のオルドビス紀中期に出現して以来、多くの形態的革新を遂げた。 初期に分岐した系統は,シルル紀からデボン紀初期にかけて,1億年以上かけて誕生した(Kenrick and Crane, 1997; Pires and Dolan, 2012; Harrison and Morris, 2018)。 胚葉類は、現存するチャロフィート群Charales、Coleochaetales、Zygnematalesに関連する淡水藻類の祖先から進化した。 蘚苔類は、胞子の発生、配偶体・胞子体の世代交代、一軸性の形態、立体的な成長など、陸上での成長への移行が観察された。 陸上維管束植物(蘚苔類、単藻類、精子類)の革新は、分岐、不定形、胞子体優勢、腋芽分岐、分裂組織・葉・根の形成を含んでいた(Harrison、2017)
上記の進化段階のいくつかは、異時性との関連が指摘されている。 胞子形成の起源については,蘚苔類のデータから,早熟な細胞質分裂,減数分裂の加速,接合体から明胞子への壁堆積の遅延を含むプロセスで接合体から直接胞子が作られたことが示唆されている(Brown and Lemmon, 2011)。 分岐胞子体(polisporangiophyte)は、頂端細胞の長大な植物成長によって進化したと仮定されている。 この植物成長の期間が長くなることで、胚軸の延長、シュートの分岐、生殖成長への移行が遅れ、胞子嚢が生成されると提唱された(Rothwell et al.2014; Tomescu et al.2014 )。 植物化石の追加研究により、古代の植物形態に関するより多くの証拠が得られ、現代と絶滅した形態の比較が可能になるはずである(Rothwell et al., 2014)。
現存植物では、配偶体発生、胚発生、植物体発生、シュート成熟、花の形態形成において異時的変化が確認されている。 Gnetumの雌性配偶体は受精と体細胞発生のタイミングの違いにより他の植物と構造的に乖離している。 受精時の精子形成や配偶子融合の時間的パターンは,細胞周期進行の時間的変化により多様化している (Friedman, 1999; Tian et al., 2005)。 子孫繁栄の場合、受精は体細胞発生の遊離核期、つまり祖先の体細胞個体発生に比べ幼い段階で起こり、卵細胞の分化が妨げられる(Friedman and Carmichael, 1998)。 Boecheraの卵巣のアポミクティックな発生は、非アポミクティック(有性)卵巣と比較して、ヘテロクロニックな遺伝子発現パターンと関連している(Sharbel et al.、2010)。 ブドウの木に感染するホロ寄生植物科のRafflesiaceaeの発生では、ネオテニーの一例と考えられる前胚期での停止と、未分化内生子から植生シュート成熟を飛ばして開花への移行が加速するという、2つのヘテロクロニックシフトが見られる(Nikolov et al, 2014)。
植生相変化が早熟なユーカリ・グロビュラスの集団と植生相変化が数年遅れる集団を比較した量的形質座位(QTL)解析では、マイクロRNA EglMIR156.5の発現がE. globulusの植生相変化の異時的変動に関係していると特定した(Hudson et al,2014)。 別のQTL解析では、Cardamine hirsutaの葉の形の自然変異の根底にヘテロクロニーがあると結論づけた(Cartolano et al.、2015)。 QTLマッピングにより、この効果は花成抑制因子ChFLCのシス制御変異によって引き起こされ、低発現のChFLC対立遺伝子を持つ集団は、早期開花と成葉形質の獲得促進、特に葉柄数の増加の両方を示すことが決定された。 形態学的およびQTL解析により、ヘテロクロニック変異がAntirrhinumの自然変異およびブドウのヘテロブラスティに寄与していることが明らかになった(Costaら、2012;Chitwoodら、2016)。 マルシダ3属(Marsilea, Regnellidium, Pilularia)の葉形の個体発生軌跡の主成分分析(PCA)により、より複雑な祖先の発生と比較して、成長速度の加速と簡略な葉形での早期終了により、ペドモフィック表現型を示すことが示された(Pryer and Hearn, 2009)。
ナス科の花序構造の進化的多様性は、花運命の獲得における異時的なシフトによって調節されている(Lippman et al, 2008; Park et al., 2012)。 家畜種と野生種のソラマメ5種のメリステム成熟のトランスクリプトームを比較したところ、発現発散のピークがあり、発育中期の発散が形態的変異を促すとする動物の胚発生に関する「逆砂時計」モデルに似ていた(Lemmon et al.、2016年)。 イネ科植物では、シュートメリステム(SM)からフローラルメリステム(FM)への移行が遅れると、より複雑なパニクルになる(Kyozuka et al.、2014)。 ポプラ(Populus sp.)やヤナギ(Salix sp.)は「カタクリ」と呼ばれるコンパクトな単性花序をつけるが、これはSMからFMへの早期移行によるパニクル形態の単純化から進化したと提案されている(Cronk et al.、2015)。 また,ヘテロクロニックな変化はMimulus guttatusの集団間における開花時期やシュート構造の自然変異にも寄与している(Baker and Diggle, 2011)。
Dipsacoideaeにおける周縁部の形態的多様性は,器官開始,特に萼の数におけるヘテロクロニック変化により引き起こされる(Naghiloo and Claßen-Bockhoff, 2017)。 アヤメ科の種間における萼片の大きな形状多様性は、ヘテロクロニー変化よりもヘテロクロニーによるものである(Guo, 2015)。 アブラナ科の研究では、多対称の祖先花から花冠の単対称性の進化には、CYC2遺伝子(TCP転写因子群)の発現が、祖先花の分裂組織での早い軸方向発現から、花弁の発生での遅い軸方向発現への異時的なシフトが関わっていることが示された(Bush et al.、2012年)。 また、ヘテロトピックではなくヘテロクロニックなCYC2発現は、ロータスにおける花弁の乳頭状円錐細胞の消失や鳥による受粉システムへの移行と関連している(Ojeda et al.、2017)。 マダガスカルツル植物(Euphorbiaceae)の一群では、花が未開花の芽のような構造で成熟花粉を保持するpaidomorphicな形態が、受粉の特殊化につながった(Armbruster et al.、2013)。 裂片状の祖先の状態から裂片状の被子植物が進化したことは、裂片状の形が幼形質を示すことから、幼形化の典型例といえる(Lord and Hill, 1987)。 キク科の裂片目は,具体的には花成の前置・後置と,雌雄異株以外のすべての胞子の新生によって進化した (Porras and Muñoz, 2000)。 ナス科のJaltomata属内の花形態の多様性は、一部の花冠形質の過形成と加速によるものである(Kostyun et al.、2017)。 また、アゾレス産の蝶形ランの多様性は、花のヘテロクロニックシフトによるものである(Bateman et al.、2014)。 最近,Ronse de Craene (2018) は,成長速度の変化と器官開始の遅延を結びつけ,フィロタクシス,共通のおしべ-花弁原基の発達,オブディプロステモニーという三つの発生過程におけるヘテロクロニーの重要性を強調した。 周縁とスタイルのヘテロクロニックな成長速度、および早期のハイパンチウムの伸長は、形態的に均質なユージニア属内の大きな種多様性の原因である(Vasconcelos et al.) 最後に、果実発生初期の細胞分裂に影響を与えるfw2.2アレルのヘテロクロニック発現は、トマト果実サイズの自然変異の原因である(Cong et al., 2002)
タイミングの遺伝子制御を解明するヘテロクロニック変異体の研究
発生タイミングに影響を与える変異体の研究は、形態形成と発生遷移の制御遺伝子経路に光を当てている。 葉状子葉(lec)変異体、ダイサー様1(dcl1)変異体、余剰子葉(xtc)変異体は、種子成熟プログラムの定義に役立つ異時的表現型を示す。lec変異体は葉と同じ特徴を持つ子葉を生み出し(Meinke, 1992)、ある構造が他の構造に置き換えられるというホメオシス現象の明らかな例である。 しかし、ホメオシスとヘテロクロニーを区別することはしばしば困難である。ホメオシスはヘテロクロニーとヘテロトピーの両方の結果である可能性があるからだ(Li and Johnston, 2000; Geuten and Coenen, 2013)。 胚発生後期において、LEC2は種子の成熟を促進し、胚発生後のアイデンティティを抑制する(Stone et al.、2008)。 dcl1変異体はlec変異体同様、胚発生期に葉緑体の発生と種子貯蔵タンパク質遺伝子の発現が野生型胚より早く起こるため、周縁型表現型を示す。 DCL1は、マスターレギュレーターであるLEC2とFUSCA3を介して種子成熟を抑制するマイクロRNAの生合成に必要である(Nodine and Bartel、2010;Willmann et al.、2011)。 xtc1、xtc2、およびanged meristem programming1 (amp1) 変異体は、最初の1〜2枚の葉が子葉に変化するホメオティック表現型を示す (Conway and Poethig, 1997)。 これらの3つの変異体では、球状体から心臓への移行が遅れ、シュートメリステムが肥大化し、胚発生時に余分な器官が形成されることになる。 この表現型は、胚の発生がより発達した形状に拡張されることから、変態の一種である超変態と解釈することができる(図1Bの変態シナリオのように)。 しかし、植物体の発生を基準に選ぶと、lec変異体は葉の同一性の獲得における前置換(peramorphosis)の場合、xtc1, xtc2, amp1は後置換(paedomorphosis)の場合に相当すると思われる。 AMP1はER膜上でのmiRNAを介した翻訳抑制に必要である(Li et al., 2013)。
植物相変化とも呼ばれる幼若期から成熟期への移行に関する遺伝子制御は、paidomorphicとperamorphic両方の変異体を用いて研究されている。 早期成体(peramorphic)表現型を示す変異体のスクリーニングでは、zippy/ago7、sgs3、rdr6などの小分子RNA生合成に関連する遺伝子の対立遺伝子が得られ( Hunter et al., 2003; Peragine et al., 2004)、一方、晩成(paidomorphic)表現型の変異体はmiRNAレベルの増加による( Gillmor et al., 2014; Xu M. et al., 2016, 2018; Guo et al., 2017 )。 これらの表現型の遺伝的基盤は、閾値依存的に、転写および翻訳の両レベルでSPL転写因子を抑制することによって作用する植物相変化の主な制御因子である、密接に関連するマイクロRNAのmiR156およびmiR157の発現変化によって説明される(Wu and Poethig, 2006; Chuck et al, 2007; Poethig, 2013; He et al.、2018)。 MIR156とSPL遺伝子ファミリーの両方は、エピジェネティックマークによって直接制御される。 MIR156は、SWR1-C複合体とクロマチンリモデラーBRAHMAによって促進されるH3K4me3とH3K4acによって活性化され、ポリコムタンパク質SWINGERとCURLY LEAFとクロマチンリモデラーPICKLEによって促進されるH3K27me3によって抑制される(Xu M. et al, 2016; Xu Y. et al., 2018; Xu M. et al, 2018)。 SPL遺伝子は、SAGA様複合体を介したヒストンアセチル化によって活性化され、ポリコムタンパク質RING1AおよびRING1Bを介したH2AUbによって抑制される(Kimら、2015;Liら、2017)。 また、SPL遺伝子の機能を喪失または獲得した植物の研究により、一部のSPL遺伝子がmiR172の発現を促進し、それがAPETALA2ファミリー開花抑制因子を抑制することで開花を促進するという内因性開花経路が定義されている(Wang et al., 2009; Wu et al., 2009; Huijser and Schmid, 2011)
大きな発生推移以外に、器官形態形成も遺伝制御因子の異時活性化によって影響を与えられることがある。 例えば、Brassica rapa胚の初期発芽は、子葉と葉の同一性がモザイク状になった器官をもたらし(Fernandez、1997)、TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATION CELL FACTOR(TCP)のクラスII遺伝子の時間差発現は、異なるサイズと形を持つ葉になる(Efroni et al, 2008)。
植物における発生タイミングの変化を駆動するメカニズム(Transcriptional, Metabolic, and Cellular Heterochrony)
転写ヘテロクロニーとは、遺伝子発現の活性化または抑制のタイミングが変わることを指し、シス制御遺伝子領域の変化によって生じることが多い(Phamら、2017)。 上記のような遺伝経路における転写ヘテロクロニーは、ヘテロクロニー表現型を生み出す一般的な方法である。 発生遷移を制御する経路の代謝制御(ここでは「代謝ヘテロクロニー」と呼ぶ)、細胞増殖・細胞拡大・細胞分化の時間制御(ここでは「細胞ヘテロクロニー」と呼ぶ)も植物におけるヘテロクロニーを駆動するメカニズムである(図2)。 転写、代謝および細胞プロセスは相互に関連しているため、ヘテロクロニーの分子的起源は複数のメカニズムの組み合わせに起因する可能性がある

FIGURE 2. 植物におけるヘテロクロニーを駆動するメカニズム。 (A)細胞内ヘテロクロニー。 植物体発生中に作られるロゼット葉の配列を示す。 シロイヌナズナコロンビア野生型植物の表現型を仮説的祖先状態とし、前置型シナリオ(ネオテニー)と周縁型シナリオ(加速)も示している。 矢印は細胞分裂停止の波の方向を示す。点線より下の細胞はまだ活発に分裂しているが、上の細胞は拡大および/または分化している。 (B) 転写と代謝のヘテロクロニー。 マイクロRNAであるmiR156/7とその標的であるSPL遺伝子の予測される存在量、および(A)の表現型に対応する植物発生の時間に沿った糖の存在量をグラフ化したものである。 点線は、幼若化(閾値以上)または成熟化(閾値未満)につながるmiR156/7とSPLの存在量の閾値を示す。 この場合、miR156の抑制とSPL発現および糖生産の活性化が遅れると幼若化し、miR156の早期の減衰とSPL発現および糖生産の早期活性化が起こると周産期化するとされている。 これらの図は簡略化した表現であり、自然界に存在する正確な量を反映したものではありません。 実際のmiR156/SPLの存在量のパターンは、「peramorphosis」状態について描かれたものに近い。
代謝異時性は、ホルモン、糖、および酸化還元シグナルによって影響を受け得る(Jia et al.、2017年)。 オーキシン、ジャスモン酸(JA)、ジベレリン、アブシジン酸などのホルモンの生合成、輸送、認識の差は、発生プロセスの制御因子を制御することでヘテロクロニーに影響を与える。 例えば、オーキシン応答因子(ARF)ARF3やARF4の変異体は成体移行を遅らせ(Fahlgren et al., 2006; Hunter et al., 2006)、オーキシンの恒常性は花幹細胞維持から雌蕊形成への移行を制御する(Yamaguchi et al., 2017)。 外因性JAはmiR156の発現低下を延期することで成体移行を遅らせ(Beydler et al., 2016)、ジベレリンはSPL遺伝子をDELLAタンパク質による抑制から解放することで開花を促進する(Yu et al., 2012)。 栄養状態は20世紀初頭から植生相変化の制御と関連している(Goebel, 1908)。 光合成によって生成される糖は成体形質の獲得に必要であり、植物体発生後期のmiR156発現の減少に一部関与している(Yang et al., 2013; Yu et al., 2013; Buendía-Monreal and Gillmor, 2017)。 HEXOKINASE1(HXK1)とトレハロース-6-リン酸(T6P)は、糖を介したmiR156の抑制に重要であり、それによって植物相変化と開花を促進する(Wahlら、2013;Yangら、2013)
細胞レベルでは器官形成は、極性の確立、細胞増殖、細胞拡張という一連の三つの段階からなる(Walcher-Chevillet and Kramer, 2016). これらのステージの開始と終了のタイミングは、器官の大きさや形状に極めて重要であり、このシーケンスの異時性により、器官の大きさや形状の多様化がもたらされる(図2)。 細胞増殖から細胞の拡大・分化への移行には、細胞周期と細胞増殖の連携が必要です(Sablowski and Carnier Dornelas, 2014)。 モデル植物の葉では、この移行は、原基の遠位部から始まり基部へと移動する細胞周期停止の基部波として移動する。 分裂停止前線の後方の細胞は高度に空胞化し、膨張を開始する(Donnelly et al., 1999; Czesnick and Lenhard, 2015)。 しかし、他の植物種では、拡散的な成長、および先端的または双方向的な細胞周期停止勾配を示すことがある(Das Gupta and Nath, 2015)。 細胞分裂から細胞拡大への移行には、光合成能力の獲得が必要である(Andriankaja et al.、2012)。 このシフトは、Target of Rapamycin (TOR) 経路を促進し、Sucrose-non-fermenting1-related kinase 1 (SnRK1) を抑制する砂糖の役割と相関している。 TORとT6Pは高分子合成を促進することで細胞拡大を誘導し、一方、SnRK1は異化を促進する(Tsai and Gazzarrini, 2014; Sablowski, 2016)。 この細胞シフトにおいて、2つのマイクロRNAが反対の役割を果たします:miR319は細胞増殖の阻害因子であるクラスII TCP因子の発現を抑制し、一方miR396は分化を遅らせるGROWTH REGULATING FACTORS (GRFs) の発現を制限します (Das Gupta and Nath, 2015; Maugarny-Calés and Laufs, 2018)。 不確定なシュート頂端分裂組織から確定的な花形分裂組織への移行には、細胞アイデンティティの時間的な制御も含まれる。 AGAMOUSがKNUCKLESを活性化し、それがWUSCHELを抑制するタイミングは、不確定性の時間窓を規定し、その結果、器官のサイズと数が決まる(Sunら、2014)
結論
植物発生の時間制御の理解は、自然界に見られる植物の形態の多様性をより良く理解し、植物形態進化を説明し、農業のために植物構造を操作するために必要となる。 上記のように、現存する植物種では、転写、代謝、細胞機構が相互に関連してヘテロクロニーを駆動しています。 今後、被子植物や基幹植物におけるこれらの経路の研究が進めば、植物の発生の進化を促した発生時期の変化について、さらに明らかになると思われる。 また、この原稿をCSGが編集・修正した。
Conflict of Interest Statement
著者らは、本研究が、利益相反の可能性があると解釈される商業的または金銭的関係のない状態で行われたことを宣言する。
Alberch, P., Gould, S., Oster, G. and Wake, D. (1979). 個体発生と系統発生における大きさと形. Paleobiology 5, 296-317. doi: 10.1017/S0094837300006588
CrossRef Full Text | Google Scholar
Andriankaja, M., Dhondt, S., DeBodt, S., Vanhaeren, H., Coppens, F., DeMilde, L.,et al. (2012). シロイヌナズナの葉の発生における増殖からの脱却:それほど漸進的ではないプロセス。 Dev. Cell 22, 64-78. doi: 10.1016/j.devcel.2011.11.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Armbruster, W. S., Lee, J., Edwards, M. E., and Baldwin, B.G. (2013). Floral paedomorphy leads to secondary specialization in pollination of Madagascar Dalechampia (Euphorbiaceae). Evolution 67, 1196-1203. doi: 10.1111/evo.12002
PubMed Abstract | CrossRef Full Text | Google Scholar
Baker, R. L., and Diggle, P. K. (2011). Am. J. Bot. 98, 1924-1934. doi: 10.3732/ajb.1100098
PubMed Abstract | CrossRef Full Text | Google Scholar
Bateman, R. M., Rudall, P. J., Bidartondo, M. I., Cozzolino, S., Tranchida-Lombardo, V., Carine, M. A.他 (2014).. アゾレス諸島の固有種チョウセンランにおける花のヘテロクロニーと推定される菌根の宿主スイッチングによる種分化. Am. J. Bot. 101, 979-1001. doi: 10.3732/ajb.1300430
PubMed Abstract | CrossRef Full Text | Google Scholar
Beydler, B. D., Osadchuk, K., Cheng, C.-L., Manak, J. R. and Irish, E.E. (2016). トウモロコシの幼若期はストレス応答遺伝子のアップレギュレーションが見られ、外来JAによって延長される。 Plant Physiol. 171, 2648-2658. doi: 10.1104/pp.15.01707
PubMed Abstract | CrossRef Full Text | Google Scholar
Brown, R. C., and Lemmon, B.E. (2011). 胞子体の前に胞子:藻類-植物移行期における胞子発生の起源を仮定する。 ニューフィトール. 190, 875-881. doi: 10.1111/j.1469-8137.2011.03709.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Buendía-Monreal, M., and Gillmor, C. S. (2017). 糖によるmiR156の収束抑制とシロイヌナズナMediatorのCDK8モジュール。 Dev. Biol. 423, 19-23. doi: 10.1016/j.ydbio.2017.01.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Busch, A., Horn, S., Mühlhausen, A., Mummenhoff, K., and Zachgo, S. (2012). 花冠の単対称性:アブラナ科における形態的新奇性の進化。 Mol. Biol. Evol. 29, 1241-1254. doi: 10.1093/molbev/msr297
PubMed Abstract | CrossRef Full Text | Google Scholar
Cartolano, M., Pieper, B., Lempe, J., Tattersall, A., Huijser, P., Tresch, A.、他 (2015).All Rights Reserved. Heterochrony underpins natural variation in Cardamine hirsuta leaf form. Proc. Natl.Acad. Sci. U.S.A. 112, 10539-10544. doi: 10.1073/pnas.1419791112
PubMed Abstract | CrossRef Full Text | Google Scholar
Chitwood, D. H., Klein, L. L., O’Hanlon, R., Chacko, S., Greg, M., Kitchen, C., et al(2016).。 ブドウの葉に埋め込まれた潜在的な発達と進化の形。 New Phytol. 210, 343-355. doi: 10.1111/nph.13754
PubMed Abstract | CrossRef Full Text | Google Scholar
Chuck, G., Cigan, A. M., Saeteurn, K., and Hake, S. (2007). トウモロコシのヘテロクロニック変異体corngrass1は、タンデム型マイクロRNAの過剰発現に起因している。 Nat. Genet. 39, 544-549. doi: 10.1038/ng2001
PubMed Abstract | CrossRef Full Text | Google Scholar
Cong, B., Liu, J., and Tanksley, S. D. (2002). トマト果実サイズ量的形質遺伝子座の自然対立遺伝子は、ヘテロクロニック調節変異によって異なる。 Proc. Natl.Acad. Sci. U.S.A. 99, 13606-13611. doi: 10.1073/pnas.172520999
PubMed Abstract | CrossRef Full Text | Google Scholar
Conway, L. J., and Poethig, R. S. (1997). 葉を子葉に変えるシロイヌナズナの突然変異。 Proc. Natl. Acad. Sci. U.S.A. 94, 10209-10214. doi: 10.1073/pnas.94.19.10209
PubMed Abstract | CrossRef Full Text | Google Scholar
Costa, M. M. R., Yang, S., Critchley, J., Feng, X., Wilson, Y., Langlade, N.、他 (2012). Antirrhinumにおけるヘテロブラスティの自然変異の遺伝的基盤。 New Phytol. 196, 1251-1259. doi: 10.1111/j.1469-8137.2012.04347.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Cronk, Q. C. B., Needham, I., and Rudall, P. J. (2015). Evolution of catkins: inflorescence morphology of selected Salicaceae in an evolutionary and developmental context(カタバミの進化:進化と発達の文脈における選択されたヤナギ科の花序の形態). Front. Plant Sci. 6:1030. doi: 10.3389/fpls.2015.01030
PubMed Abstract | CrossRef Full Text | Google Scholar
Czesnick, H., and Lenhard, M. (2015). 植物におけるサイズコントロール-葉と花からの教訓. コールドスプリングハーブ. Perspect. Biol. 7:a019190. doi: 10.1101/cshperspect.a019190
PubMed Abstract | CrossRef Full Text | Google Scholar
Das Gupta, M, and Nath, U. (2015). 葉の成長極性のパターンのダイバージェンスは、miR396の発現ダイバージェンスと関連している。 Plant Cell 27, 2785-2799. doi: 10.1105/tpc.15.00196
PubMed Abstract | CrossRef Full Text | Google Scholar
De Beer, G. R. (1951). 胚と祖先. オックスフォード: Clarendon Press.
Google Scholar
Donnelly, P. M., Bonetta, D., Tskaya, H., Dengler, R. E., and Dengler, N. G. (1999)(1999). シロイヌナズナの発育葉における細胞周期と細胞の肥大化。 Dev. 215, 407-419. doi: 10.1006/dbio.1999.9443
PubMed Abstract | CrossRef Full Text | Google Scholar
Efroni, I., Blum, E. , Goldshmidt, A. and Eshed, Y. (2008)(2018. シロイヌナズナの葉の発生には、長期的かつ動的な成熟スケジュールが存在する。 また、このような研究成果をもとに、「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「植物細胞」「微生物」「植物細胞」「微生物」「微生物」「微生物」「植物細胞」「微生物」「植物細胞」「微生物」「微生物」「植物細胞」「植物細胞」「植物細胞」「植物細胞 TAS3 ta-siRNAによるAUXIN RESPONSE FACTOR3の制御は、シロイヌナズナの発生タイミングとパターニングに影響を及ぼす。 Curr. 16, 939-944. doi: 10.1016/j.cub.2006.03.065
PubMed Abstract | CrossRef Full Text | Google Scholar
Fernandez, D. E. (1997). アブラナ科植物の早発芽胚におけるホメオシスの発生的基盤:シュート頂点の相変化。 Development 124, 1149-1157.
PubMed Abstract | Google Scholar
Friedman, W. E. (1999). シロイヌナズナの精子における細胞周期の発現:植物および他の真核生物における配偶子形成および受精のパターンの理解への示唆。 Development 126, 1065-1075.
PubMed Abstract | Google Scholar
Friedman, W. E., and Carmichael, J. S. (1998)(2010). ヘテロクロニーと発生革新:高度にApomorphicな種子植物であるgnetumにおける雌性配偶体の個体発生過程の進化。 Evolution 52, 1016-1030. doi: 10.1111/j.1558-5646.1998.tb01830.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Geuten, K., and Coenen, H. (2013). 植物の進化と発生におけるヘテロクロニック遺伝子. Front. Plant Sci. 4:381. doi: 10.3389/fpls.2013.00381
PubMed Abstract | CrossRef Full Text | Google Scholar
Gillmor, C. S., Silva-Ortega, C. O., Willmann, M. R., Buendía-Monreal, M., and Poethig, R. S. (2014). シロイヌナズナのメディエーターCDK8モジュール遺伝子CCT(MED12)およびGCT(MED13)は、発生段階遷移のグローバルレギュレーターである。 Development 141, 4580-4589. doi: 10.1242/dev.111229
PubMed Abstract | CrossRef Full Text | Google Scholar
Goebel, K. (1908). 植物実験形態学入門. Leipzig: B.G. Teubner.
Google Scholar
Gould, S. J. (1977). Ontogeny and Phylogeny. Cambridge, MA: Harvard University Press.
Google Scholar
Guo, C., Xu, Y., Shi, M., Lai, Y., Wu, X., Wang, H., et al. (2017). miR159によるmiR156の抑制は、シロイヌナズナの幼若期から成熟期への移行時期を制御している。 Plant Cell 29, 1293-1304. doi: 10.1105/tpc.16.00975
PubMed Abstract | CrossRef Full Text | Google Scholar
Guo, J. (2015). 発生配列の比較解析により明らかになったアヤメの複雑で多様に精巧な萼片の進化機構。 Am. J. Bot. 102, 819-832. doi: 10.3732/ajb.1400519
PubMed Abstract | CrossRef Full Text | Google Scholar
Haeckel、E. (1875).。 Die Gastrula und die Eifurchung der Thiere. Jena. Z. Naturwiss. 9, 402-508.
Google Scholar
Harrison, C. J. (2017)(2017). 陸上植物のボディプランの進化における発達と遺伝学. Philos. Trans. R. Soc. B Biol. Sci. 372:20150490. doi: 10.1098/rstb.2015.0490
PubMed Abstract | Cross Ref Full Text | Google Scholar
Harrison, C.J., and Morris, J.L. (2018). 維管束植物の新芽と葉の起源と初期進化. Philos. Trans. R. Soc. B Biol. Sci. 373:20160496. doi: 10.1098/rstb.2016.0496
PubMed Abstract | Cross Ref Full Text | Google Scholar
He, J., Xu, M., Willmann, M. R., Mccormick, K., Hu, T., Yang, L., et al. (2018). miR156/miR157によるSPL遺伝子発現の閾値依存的抑制は、シロイヌナズナの植生相変化を制御する。 PLoS Genet. 14:e1007337. doi: 10.1371/journal.pgen.1007337
PubMed Abstract | CrossRef Full Text | Google Scholar
Hudson, C. J., Freeman, J. S., Jones, R. C., Potts, B. M., Wong, M. M. L., Weller, J. L., et al (2014). ユーカリ・グロブルスにおけるヘテロクロニーの遺伝的制御. G3 4, 1235-1245. doi: 10.1534/g3.114.011916
PubMed Abstract | CrossRef Full Text | Google Scholar
Huijser, P., and Schmid, M. (2011). 植物における発生段階遷移の制御. Development 138, 4117-4129. doi: 10.1242/dev.063511
PubMed Abstract | CrossRef Full Text | Google Scholar
Hunter, C., Sun, H. and Poethig, R. S. (2003). シロイヌナズナのヘテロクロニック遺伝子ZIPPYはARGONAUTEファミリーの一員である。 Curr. Biol. 13, 1734-1739. doi: 10.1016/j.cub.2003.09.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Hunter, C., Willmann, M. R., Wu, G., Yoshikawa, M., de la Luz Gutiérz-Nava, M. and Poethig, R. S. (2006). ETTINとARF4のsiRNAによる抑制は、シロイヌナズナのヘテロブラスティを制御する。 Development 133, 2973-2981. doi: 10.1242/dev.02491
PubMed Abstract | CrossRef Full Text | Google Scholar
Jia,X. L., Chen,Y. K, Xu, X. Z., Shen, F., Zheng, Q. B., Du, Z., et al. (2017). miR156 switches on vegetative phase change under the regulation of redox signals in apple seedlings.リンゴ苗の酸化還元シグナルの制御下で植物相の変化を促す。 Sci. Rep. 7, 1-13. doi: 10.1038/s41598-017-14671-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Kenrick, P. and Crane, P. R. (1997). 陸上植物の起源と初期進化(The origin and early evolution of plants on land). Nature 389, 33-39. doi: 10.1038/37918
CrossRef Full Text | Google Scholar
Keyte, A. L., and Smith, K.K. (2014). ヘテロクロニーと発生タイミング機構:進化におけるオントジェニーの変化. Semin. Cell Dev. Biol. 34, 99-107. doi: 10.1016/j.semcdb.2014.06.015
PubMed Abstract | CrossRef Full Text | Google Scholar
Kim, J.Y., Oh, J. E., Noh, Y.S., and Noh, B. (2015). シロイヌナズナSAGA様複合体による幼若期から成熟期への移行をエピジェネティックに制御する。 Plant J. 83, 537-545. doi: 10.1111/tpj.12908
PubMed Abstract | CrossRef Full Text | Google Scholar
Kostyun, J. L., Preston, J. C., and Moyle, L. C.(2017).シロイヌナズナ(幼植物から成植物への移行)。 Heterochronic developmental shifts underlie floral diversity within Jaltomata ( Solanaceae ). Evodevo 8:17. doi: 10.1186/s13227-017-0080-z
PubMed Abstract | CrossRef Full Text | Google Scholar
Kyozuka, J., Tokunaga, H. and Yoshida, A. (2014). メリステムの相変化の微調整によるイネ科花序の形状の制御. Curr. Opin. Plant Biol. 17, 110-115. doi: 10.1016/j.pbi.2013.11.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Lemmon, Z. H., Park, S. J., Jiang, K., Van Eck, J., Schatz, M. C., and Lippman, Z. B. (2016). ナイトシェードにおける花序の多様性の進化と分裂組織成熟時のヘテロクロニー。 Genome Res. 26, 1676-1686. doi: 10.1101/gr.207837.116
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, J., Wang, Z., Hu, Y., Cao, Y. and Ma, L.(2017).[邦訳]斉藤茂樹、斉藤茂樹、斉藤茂樹、斉藤茂樹、斉藤茂樹(2017). ポリコームグループタンパク質RING1AとRING1Bはシロイヌナズナの植物相転移を制御している。 Front. Plant Sci. 8:867. doi: 10.3389/fpls.2017.00867
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, P., and Johnston, M. O. (2000). 20世紀の植物進化研究におけるヘテロクロニー(Heterochrony). Bot. Rev. 66, 57-88. doi: 10.1007/BF02857782
CrossRef Full Text | Google Scholar
Li, S., Liu, L., Zhuang, X., Yu, Y., Liu, X., Cui, X, and al. (2013). マイクロRNAはシロイヌナズナの小胞体上の標的mRNAの翻訳を阻害する。 Cell 153, 562-574. doi: 10.1016/j.cell.2013.04.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Lippman, Z. B., Cohen, O., Alvarez, J. P., Abu-Abied, M., Pekker, I., Paran, I.、その他 (2008). トマトと関連するナイトシェードにおける複散形花序の形成。 PLoS Biol. 6:e288. doi: 10.1371/journal.pbio.0060288
PubMed Abstract | CrossRef Full Text | Google Scholar
Lord, E. M., and Hill, J. P. (1987). “Evidence for heterochrony in the evolution of plant form,” in Development as an Evolutionary Process, eds R. A. Raff and E. C. Raff (ニューヨーク、NY. Alan R. Liss), 47-70.
グーグルスカラー
Maugarny-Calés, A. and Laufs, P. (2018年). 葉を形にする:分子、細胞、環境、進化的な視点。 Development 145:dev161646. doi: 10.1242/dev.161646
PubMed Abstract | CrossRef Full Text | Google Scholar
Meinke,D. W. (1992). 葉状子葉を持つA homoetic mutant of Arabidopsis thaliana. Science 258, 1647-1649. doi: 10.1126/science.258.5088.1647
PubMed Abstract | CrossRef Full Text | Google Scholar
Naghiloo, S., and Claßen-Bockhoff, R. (2017)(2019. 時間と空間における発生的変化が花の進化的多様化を促進する:Dipsacoideaeのケーススタディ。 Front. Plant Sci. 8:1665. doi: 10.3389/fpls.2017.01665
PubMed Abstract | CrossRef Full Text | Google Scholar
Nikolov, L. A., Tomlinson, P. B., Manickam, S., Endress, P. K., Kramer, E. M., and Davis, C. C. (2014). ホロホロチョウセンアサガオは最も減少したエンドファイトを持ちながら、世界最大の花を生み出す。 Ann. Bot. 114, 233-242. doi: 10.1093/aob/mcu114
PubMed Abstract | CrossRef Full Text | Google Scholar
Nodine, M. D., and Bartel, D.P. (2010). マイクロRNAは植物の胚発生において早熟な遺伝子発現を防ぎ、パターン形成を可能にする。 Genes Dev. 24, 2678-2692. doi: 10.1101/gad.1986710
PubMed Abstract | CrossRef Full Text | Google Scholar
Ojeda, D. I., Jaén-Molina, R., Santos-Guerra, A., Caujape-Castells, J. and Cronk, Q. (2017).植物胚発生の際に、早熟な遺伝子発現を防ぎ、パターン形成を可能にする。 花弁アイデンティティ遺伝子の発現パターンの時間的、しかし空間的な変化は、マカロニハス(マメ科)における乳頭状円錐細胞の消失と鳥による受粉への移行と関連している。 Plant Biol. 19, 420-427. doi: 10.1111/plb.12551
PubMed Abstract | CrossRef Full Text | Google Scholar
Park, S. J., Jiang, K., Schatz, M. C., and Lippman, Z. B. (2012). トマトにおける分裂組織の成熟速度が花序の構造を決定する。 Proc. Natl. Acad. Sci. U.S.A. 109, 639-644. doi: 10.1073/pnas.1114963109
PubMed Abstract | CrossRef Full Text | Google Scholar
Peragine, A., Yoshikawa, M., Wu, G., Albrecht, H. L. and Poethig, R. S. (2004). SGS3とSGS2/SDE1/RDR6はシロイヌナズナの幼若期発生とトランス作用型si RNAの産生に必要である。 Genes Dev. 18, 2368-2379. doi: 10.1101/gad.1231804.nism
CrossRef Full Text | Google Scholar
Pham, T., Day, S. M., Glassford, W. J., Williams, T. M., and Rebeiz, M. (2017)(2019. 極端なヘテロクロニックシフトによる新規発現パターンの進化的起源. Evol. Dev. 19, 43-55. doi: 10.1111/ede.12215
PubMed Abstract |Ref Full Text | Google Scholar
Pires, N.D., and Dolan, L. (2012). 陸上植物における形態進化:古い遺伝子による新しいデザイン. Philos. Trans. R. Soc. B Biol. Sci. 367, 508-518. doi: 10.1098/rstb.2011.0252
PubMed Abstract | Cross Full Text | Google Scholar
Poethig, R. S. (2013). 植物における植生相変化とシュート成熟、第1版。 New York, NY: Elsevier Inc. doi: 10.1016/B978-0-12-396968-2.00005-1
PubMed Abstract | CrossRef Full Text | Google Scholar
Porras, R., and Muñoz, J. M. (2000)(2010. キク科の花被片:異時的起源。 Am. J. Bot. 87, 925-933. doi: 10.2307/2656991
PubMed Abstract | CrossRef Full Text | Google Scholar
Pryer, K. M., and Hearn, D. J. (2009).を参考にした。 このような状況下において、「萌芽的」であることが重要である。 Evolution 63, 498-513. doi: 10.1111/j.1558-5646.2008.00562.x
PubMed Abstract | Cross Full Text | Google Scholar
Ronse de Craene, L. (2018). 被子植物の花の進化における花成の役割の理解:歴史的・物理力学的観点からの解明。 J. Plant Res. 131, 367-393. doi: 10.1007/s10265-018-1021-1
PubMed Abstract |Ref Full Text | Google Scholar
Rothwell, G. W., Wyatt, S. E., and Tomescu, A. M. F. (2014). 古生物学と発生生物学の接点における植物進化:生物中心のパラダイム。 Am. J. Bot. 101, 899-913. doi: 10.3732/ajb.1300451
PubMed Abstract | CrossRef Full Text | Google Scholar
Sablowski, R.(2016). 植物細胞の成長・分裂の調整:集団制御か相互合意か? Curr. Opin. Plant Biol. 34, 54-60. doi: 10.1016/j.pbi.2016.09.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Sablowski, R., and Carnier Dornelas, M. (2014). 植物における細胞増殖と細胞周期の相互作用. J. Exp. Bot. 65, 2703-2714. doi: 10.1093/jxb/ert354
PubMed Abstract | CrossRef Full Text | Google Scholar
Sharbel, T. F., Voigt, M. L., Corral, J. M., Galla, G., Kumlehn, J., Klukas, C.,et al. (2010). ボエケラのアポミクティックおよび有性卵巣は、ヘテロクロニックなグローバル遺伝子発現パターンを示す。 Plant Cell 22, 655-671. doi: 10.1105/tpc.109.072223
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, K.K. (2002). 配列の異時性(heterochrony)と発生の進化(evolution of development)。 J. Morphol. 252, 82-97.10014
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, K. K. (2003). 時間の矢:ヘテロクロニーと発達の進化. Int. J. Dev. Biol. 47, 613-621.
Google Scholar
Stone, S. L., Braybrook, S. A., Paula, S. L., Kwong, L. W., Meuser, J., Pelletier, J., and al. (2008). シロイヌナズナ葉状子葉2は成熟形質とオーキシン活性を誘導する:体細胞胚形成への示唆。 Proc. Natl.Acad. Sci. U.S.A. 105, 3151-3156. doi: 10.1073/pnas.0712364105
PubMed Abstract | CrossRef Full Text | Google Scholar
Sun, B., Looi, L. S., Guo, S., He, Z., Gang, E. S., Huang, J., et al(2014).シロイノシシの成熟形質とオーキシンの活性を誘導し、体細胞胚発生を示唆する。 植物幹細胞におけるPolycomb evictionによって細胞分裂に依存したタイミング機構が発動される。 Science 343:1248559. doi: 10.1126/science.1248559
PubMed Abstract | CrossRef Full Text | Google Scholar
Tian, H. Q., Yuan, T., and Russell, S. D. (2005). タバコの雄・雌の配偶子における二重受精と細胞周期の関係. Sex. Plant Reprod. 17, 243-252. doi: 10.1007/s00497-004-0233-9
CrossRef Full Text | Google Scholar
Tomescu, A. M. F., Wyatt, S. E., Hasebe, M., and Rothwell, G. W. (2014). 維管束植物のボディプランの初期進化-失われたメカニズム-. Curr. Opin. Plant Biol. 17, 126-136. doi: 10.1016/j.pbi.2013.11.016
PubMed Abstract | CrossRef Full Text | Google Scholar
Tsai, A.Y.-L., and Gazzarrini, S. (2014). 植物の発生とシグナル伝達におけるトレハロース-6-リン酸とSnRK1キナーゼ:新たな絵。 Front. Plant Sci. 5:119. doi: 10.3389/fpls.2014.00119
PubMed Abstract | CrossRef Full Text | Google Scholar
Vasconcelos, T. N. C., Lucas, E. J., Faria, J. E. Q., and Prenner, G. (2018). Floral heterochrony promotes flexibility of reproductive strategies in the morphologically homogeneous genus Eugenia (Myrtaceae). Ann. Bot. 121, 161-174. doi: 10.1093/aob/mcx142
PubMed Abstract | CrossRef Full Text | Google Scholar
Wahl, V., Ponnu, J., Schlereth, A., Arrivault, S., Langenecker, T., Franke, A.ほか (2013).Onc. シロイヌナズナにおけるトレハロース-6-リン酸シグナルによる開花の制御(Regulation of flowering by trehalose-6-phosphate signaling in Arabidopsis thaliana. Science 339, 704-707. doi: 10.1126/science.1230406
PubMed Abstract | CrossRef Full Text | Google Scholar
Walcher-Chevillet, C. L., and Kramer, E. M. (2016). Breaking the mold: Understanding the evolution and development of lateral organs in diverse plant models(型にはまらない:多様な植物モデルにおける側器官の進化と発達の理解). Curr. Opin. Genet. Dev. 39, 79-84. doi: 10.1016/j.gde.2016.06.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Wang, J., Czech, B., and Weigel, D. (2009). miR156-制御SPL転写因子によりシロイヌナズナの内因性の開花経路を定義する. Cell 138, 738-749. doi: 10.1016/j.cell.2009.06.014
PubMed Abstract | CrossRef Full Text | Google Scholar
Willmann, M. R., Mehalick, A. J., Packer, R. L., and Jenik, P.D. (2011). マイクロRNAはシロイヌナズナの胚成熟のタイミングを制御している。 Plant Physiol. 155, 1871-1884. doi: 10.1104/pp.110.171355
PubMed Abstract | CrossRef Full Text | Google Scholar
Wu, G., Park, M.Y., Conway, S.R., Wang, J. , Weigel, D., and Poethig, R. S. (2009年). miR156とmiR172の連続的な作用は、シロイヌナズナの発生タイミングを制御している。 Cell 138, 750-759. doi: 10.1016/j.cell.2009.06.031
PubMed Abstract | CrossRef Full Text | Google Scholar
Wu, G.and Poethig, R. S. (2006). シロイヌナズナにおけるmiR156とその標的SPL3によるシュート発生の時間的制御。 Development 133, 3539-3547. doi: 10.1242/dev.02521
PubMed Abstract | CrossRef Full Text | Google Scholar
Xu, M., Hu, T., Smith, M. R. , and Poethig, R.S.(2016).シロイヌナズナにおけるシュート発生の時間的制御。 シロイヌナズナにおける植生相変化のエピジェネティック制御 Plant Cell 28, 28-41. doi: 10.1105/tpc.15.00854
PubMed Abstract | CrossRef Full Text | Google Scholar
Xu, Y., Guo, C. , Zhou, B. , Li, C. , Wang, H. , Zheng, B., et al(2016)。 SWI2/SNF2クロマチンリモデリングATPase BRAHMAによる植生相変化の制御. Plant Physiol. 172, 2416-2428. doi: 10.1104/pp.16.01588
PubMed Abstract | CrossRef Full Text | Google Scholar
Xu, M., Leichty, A. R., Hu, T. and Poethig, R. S. (2018). H2A.ZはH3K4me3の沈着を促進することで、シロイヌナズナのMIR156AおよびMIR156Cの転写を促進する。 Development 145:dev.152868. doi: 10.1242/dev.152868
PubMed Abstract | CrossRef Full Text | Google Scholar
Yamaguchi, N., Huang, J., Xu, Y., Tanoi, K. and Ito, T.(2017).シロイヌナズナのMIR156AとMIR156Cの転写を促進するH2A.Z。 オーキシンのホメオスタシスの微調整は、花幹細胞維持から雌性体形成への移行を支配している。 Nat. Commun. 8:1125. doi: 10.1038/s41467-017-01252-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Yang, L., Xu, M., Koo, Y., He, J. and Poethig, R. S.(2013). 糖はMIR156AとMIR156Cの発現を抑制することでシロイヌナズナの植物相変化を促進する.eLife 2:e00260.
PubMed Abstract | CrossRef Full Text | Google Scholar
Yu, S., Li, C., Zhou, C. M., Zhang, T. Q., Lian, H., Sun, Y., et al. Sugar is an endogenous cue for juvenile-to-adult phase transition in plants. eLife 2:e00269. doi: 10.7554/eLife.00269
PubMed Abstract | CrossRef Full Text | Google Scholar
Yu, S., Zhang, Y., Horrer, D., Zhang, T. , Hao, Y., Feng, Y. , et al. ジベレリンはmiR156を標的としたSQUAMOSAプロモーター結合-様転写因子を介してシロイヌナズナの花成変化を制御する. Plant Cell 24, 3320-3332. doi: 10.1105/tpc.112.101014
PubMed Abstract | CrossRef Full Text | Google Scholar
.
Leave a Reply