Anatomy of Cardiac Nodes and Atrioventricular Specialized Conduction System|Revista Española de Cardiología
INTRODUCTION
1852年のStannius1 の古典的研究は、心伝導が筋原性であると最初に提案したものである。 約 1 世紀前、特殊な筋肉組織が心拍の開始と伝播に関与していることが示されました。 1906年、俵直2が、1893年にHis3によって報告された筋束の存在を確認した。 さらに1906年、KeithとFlack4は、His-Tawaraシステムの存在を確認する。 その1年後、彼らは中耳(SA)結節の構造を説明した5
プルキンエ6が初めて特殊な心室繊維を記述したが、彼は心臓の構造におけるその重要性に気づいておらず、ヒスの記述した筋束が心室プルキンエ繊維と連続していることを示したのは俵2である6。 俵の研究は最近になって英語に翻訳された7 が、彼の研究の一部を初めて英語に翻訳したのは、Robb8 が1965年に出版した教科書であった。 これらの著作は、心伝導系を研究しようとするすべての研究者にとって基本的な読み物である。
COMPONENTS OF THE SPECIFIC ATRIOVENTRICULAR CONDUCTION SYSTEM
心房から心室につながる筋束は His3 によって “penetrating bundle” と表現された。 しかし、Hisはこの筋束が右心房で房室結節、心室、心室プルキンエ細胞と連続していることを組織学的に観察していない。 俵2が、アショフの指導のもとに博士号を取得する過程で、この結合を認識したのである。 10 その後、これらの結合は健康な心臓には見られず、病気の心臓に見られることが確認された。 その結果、上大静脈と右心房が結合する部分にあることが疑われた。実験条件下では、この部分は心臓の中で最後に鼓動を停止する部分(いわゆるultimum moriens)となっていたのである。 1907年、KeithとFlack5 は、ヒトを含むすべての哺乳類において、SA結節または洞結節を識別した。 その構成細胞は心臓インパルスの発生部位であると考えられた。
CSは右心房の前上部にあるSAノードで発生する(図1)。 房室結節は心房内の下部、後部の位置にある。 房室結節から心房細動を経て、左右の束枝に分岐し、結合組織の鞘に包まれて周囲の筋組織から隔離されながら、心室間中隔を下降する。 心筋内ではプルキンエ回路網と連続している(図1)
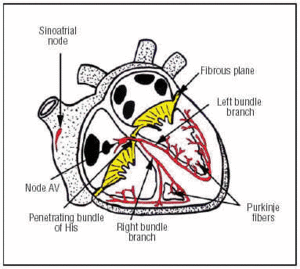
Fig. 心伝導系を図式化したもの(赤色)。
形態学的-巨視的関心領域
いくつかの巨視的関心領域は、心臓CSの位置を特定するのに役立つ。 SA結節は心外膜下にあり(図2a)、上大静脈の筋組織と心房付属器の筋組織の間の接合部に挟まれている。 その基部は末端紋章の反対側にある。 SA結節と心外膜との距離は0.3±0.1mmである。11 約10%の人では、結節は下大静脈の方に伸びず、上大静脈の開口部の下部の周りに馬蹄形になっている12。 房室結節は、Kochが最初に描いた三角形の領域の頂点にある心房中隔の基部に存在する13。この三角形は右心房の心内膜表面にあり(図2b、c)、前方は三尖弁の中隔葉の挿入部で、後方はTodaroの腱として知られる線維性の腱で縁取られている。 この腱は耳小骨弁の心内膜下の繊維状の部分で、冠状静脈洞の開口部と卵円窩を隔てる心房筋に挿入される。 この三角形の頂点は、上記の前縁と後縁の接合部によって上方に形成されており、心臓の中心線維体(CFB)に相当する。 三角形の底辺は冠状静脈洞の開口部と三尖弁の中隔リーフレットを支える右心房の前庭で形成されている。 この底部は電気生理研究者の間では中隔峡と呼ばれ、房室結節リエントリー性頻拍の患者にはここで徐脈路の高周波アブレーションが行われます14
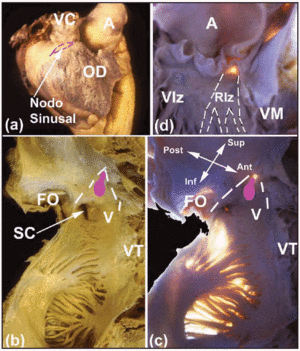
図2. A:右心房の外側心外観。SAノードの位置はピンクの破線で示す。 BおよびC:右心房の後壁および中隔壁の心内膜像(通常および透過照明あり)で、卵円窩(OF)およびKochの三角形の限界(白の破線)、トダゾーの腱(TT)および三尖弁(TV)の中隔尖の挿入を示す。 右心房の前庭(V)と冠状静脈洞の開口部(CS)が下限を形成している。 房室結節の位置は楕円形の結節(ピンク色)で示す。 D:膜状中隔を示す左心室像(透過照明)。 ここがHis束の出現点であり、右束枝、左束枝と連続する。 左束枝(LBB)は白の破線で示した。 Aは大動脈、AAは心房付属器、VCは上大静脈、RVは右心室、LVは左心室、MVは僧帽弁を示す。
AV伝導の継続は、CFBを貫く伝導軸の唯一の部分であるHis束の貫通を介して起こる。 CFBは大動脈弁および僧帽弁の葉の結合組織と三尖弁の中隔葉(いわゆる右線維性三尖)および心室間中隔の膜状部分との結合により形成されている。 多くの哺乳類の心臓では三尖は線維性であるが、ウシの心臓は中央に骨や軟骨の塊(索状突起)がある。 一方、マッコウクジラのCFBは繊維組織が非常に緩やかである。 膜状部分または隔壁は、長さに幅があるが、AV束Hisを見つけるための良い指針となる。 これは右線維性三角錐を横切った後、この膜状部分の上に現れ(図2d)、その後左右の束枝に分かれる。 右枝は右心室内側乳頭筋の基部で中隔筋を通過する。 その後、細い索となり、内側乳頭筋と前乳頭筋をつなぐ中膜海綿体またはモデレータ帯の奥深くまで入り込みます。 左枝の起始部は大動脈弁の右尖と非冠尖の交叉部の下にあり、その後、心室中隔の心内膜下を下降する(図2d)。 その経路は、光沢のある繊維状の薄板で覆われているため、時々見えることがある。 左枝の近位部は右枝よりはるかに長い。 胎児や乳児の心臓では、時に「行き止まり路」15と呼ばれる第3の枝が見られることがあるが、これは大動脈の根元に向かって前上方方向に束縛が続いているものである。
結節の構造と房室伝導系
俵2やその後のDavies16、Truexら17(ほんの一例)のような組織学的手法を用いた研究により、ヒトの房室は他の哺乳類のものとよく似た配列になっている(種や心臓によって多少の違いはある)ことが示されている。 俵2 は、特殊化した心筋細胞が、光学顕微鏡で見える薄い結合組織のシートによって、正常心筋細胞や作業心筋細胞と分離していることを報告し、これに基づいて、Aschoff18 と Mönckeberg19 が特殊心筋の組織学的識別のために提案した基準となっている。 簡単に言えば、特殊化した心筋細胞は、光学顕微鏡で見たときに作業筋細胞よりも目立ち、ある組織切片から次の組織切片へと「追跡」することができるのである。 Robb8はその単行本の中で、導電性組織を「導電系」ではなく「接続系」という言葉で定義することを好んだ。なぜなら、組織学的標本は機能よりも細胞の形態をよく定義してくれるからである。 彼はまた、剖検材料の鮮度や使用した固定・染色法によって、特殊な心筋の質感に違いがあることを観察している。 俵2もこのことに気づいており、心臓の組織切片でも特殊化した筋細胞の形態が不均一であることを指摘している。 近年、ヒトや他の哺乳類の胎生期の心臓の導電性組織の位置を特定するために、さまざまな分子マーカーや免疫組織化学マーカーが使用されている。
正常なヒトの心臓において、SAおよびAV結節は、結合組織または脂肪組織によって周囲の心筋から電気的に絶縁されていないため、AschoffおよびMönckebergの基準18、19を満たさない。 むしろ、移行細胞からなる小さな領域を経て、心房の作業線維と接触している。 SAノードでは、KeithとFlack5が洞細胞と作動細胞を区別している。 しかし、俵2 は房室結節とHis束の細胞との鑑別に困難があることを指摘している。 しかし、俵2 は、房室結節とHis束の細胞との区別が難しいことを指摘し、両者の違いは純粋に解剖学的なものであることを提唱した。 この定義に基づき、CSのCFBに完全に覆われた部分をpenetrating bundleまたはbundle of Hisと呼ぶ(図3a)。 また、近位伝導系からHis束までの心房部分は房室結節と呼ばれる(図3b)。 この解剖学的区別は、貫通束が絶縁体であるため、求心性心房の電気的活動に直接接触することができないことから論理的なものである。 したがって、心房の活動はすべて房室結節を経由して事前に指示されなければならない。 van Gieson法で染色したヒト(a;10倍)とブタ(b;40倍)の心臓の洞房(SA)結節の矢状断面組織図。 洞房細胞(SC)と作業心房細胞(WAC)の接触に注意。 洞房細胞はより明瞭で、より多くの結合組織(赤色)に埋め込まれているのが特徴。 c: van Gieson法によるKochの三角形の中央部の染色。 コンパクトAVノードの形状とコンパクトノードの凸面に接する移行細胞(TC)に注目。 d: d:CFBからの線維組織(緑)に囲まれたHisの貫通束を示すMassonの三色染色された切片。 SNAは洞結節動脈,CFBは中心線維体,TVは三尖弁を示す。
SA結節の本来の機能は心臓インパルスの源となることである。 ヒトのSA結節は弓状または瘤状の構造をしている。 組織学的には、通常の作業細胞よりわずかに小さい細胞が束になって配置されている。 これらは空間的な秩序なく混在し、染色性は弱く、高密度の結合組織マトリックスに埋め込まれている(図3 aおよびb)。 21 結節の周辺部では、特殊な細胞が作業筋の細胞と混在している(図3a、b)。 さらに、作業心筋と相互接続する複数の放射状または伸展部が報告されている。 これらは心筋内を通り、末端堤防や上・下大静脈に入り込んでいる。 SA結節は洞結節動脈と呼ばれる動脈の周囲に配置されており、この動脈は結節内の中心または偏心に走ることができる。 ヒトの心臓の29%において、この動脈は結節の内部を走行している。11 SA結節はまた、自律神経系と密接に関連している。 これらの神経線維の大部分は副交感神経であり、交感神経線維は結節の血管の周りに集中していることが示唆されている23
房室の固有の機能は心臓インパルスを遅延させることである。 ヒトの場合、この結節はコンパクトな部分と過渡的な細胞の領域がある。 前者は半楕円形でCFBの上に位置している(図3c)。 Kochの三角形の底に近い部分では、房室結節のコンパクトな部分は2つの拡張部または伸張部に分かれている。 房室結節に血管を張り巡らす動脈は通常これらの間に存在する。 24 移行細胞の大きさは、房室結節と心房の作業細胞の中間である。 これらの細胞は、作業細胞よりも多くの結合細胞に囲まれているが、隣接する心筋から絶縁されてはいない。 しかし、SA結節からのインパルスがどのようにして房室結節に到達するかについては議論が分かれるところである。 26
房室結節は遠位で貫通するHis束と続いているが(図3d)、His束の細胞がより平行に配置されているなど、この2つの構造には細胞の配置の点で若干の違いがある。
イヌの房室結節はヒトのそれよりも小さいが、貫通したHis束が長い27。 ウサギでは、His束の一部が房室結節の一部であるかのように記述されているが29、これは誤りである(図4a〜d)。 イヌの房室結節とウサギやヒトの房室結節との最も顕著な形態的差異は、前者が移行性細胞で覆われていないことである。 ラット(安静時の心拍数はイヌやヒトの10倍)では、房室結節はイヌのそれと比例しているが、CFBは小さい。
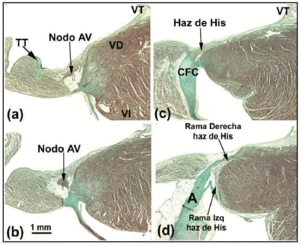
図4. この合成図は、ウサギの房室結節とHisバンドルとその左右の束枝を示したものである。 bの横棒は1mmを表す(すべての画像で同じ)。 Masson’s trichrome染色。
伝導系の組織学的軌跡を貫通したヒス束に向かってたどると、多くのヒトの心臓でヒス束は左向きに曲がり、間中隔の筋稜上に出ていることがわかる。 この束はCFBからの結合組織に取り囲まれており、左右の束に分かれるまでの長さは様々である。 前者は心室間中隔の左側で分岐する(図5a、c)。 束束の分岐は、心室間中隔の筋紋の上にしゃがんでいる騎手のような形をしています(図5a)。 しかし、時には左側に偏位していることもある(図5c)。 このような場合、右枝は中隔の筋組織の内部に入り(図5b)、内側乳頭筋の挿入に関連して右心室内に現れる。 b:この心臓の右枝はその起始部で心筋内にあり,結合組織(青)に囲まれている(Jones’ trichrome stain, x20)。 c:この心臓のHis束の分岐は、心室間中隔の筋紋の左側にずれており、左心室の心内膜の下で縦方向に下降している(Jones´thichrome、x5)。 d:左枝の被覆結合鞘(青)に注目(Jones´thichrome、x10)。 Aは大動脈、Eは心内膜、TVは三尖弁。
その近位部に沿って、右および左束枝は線維性薄板で覆われている(図5bおよびd)。 俵2が示したように(図6a)、ヒトの場合、左枝は通常3つの束に分かれ、広範囲に渡って相互に連絡を取っている。 これらの束は心尖部でramifiedとなり、僧帽弁の2つの乳頭筋の内部まで伸びているが、心室壁に沿って心基部まで戻っている。 さらに遠くのヒトの心室頂部では、プルキンエ線維は繊維状の被膜を失い、働く心筋と同じように見えるので、その稜線をたどることはほとんど不可能になる。
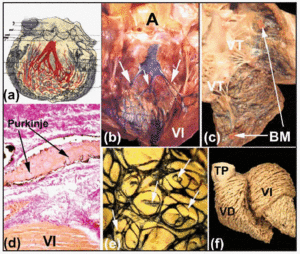
図6 a:ヒトの左束枝の三叉配置を示す俵の図 bとc:右心室と左心室を開いた新しい子牛心臓。 心内膜下に墨汁を注入すると、左右の束枝とプルキンエ網が見える。 d:子牛の心臓の切片(van Gieson染色、100倍)で、プルキンエネットワークにイン ドインクを注入した後、結合組織(赤)でその端を包んでいる。 ネットワークの楕円形の配置と、端から心筋を貫く分枝(矢印)に注目。 f:ヒト成人心臓の心室筋線維の解剖。 左心室の内層と深層の配置の違いに注意。 Aは大動脈、PTは肺動脈、LVは左心室、TVは三尖弁を示す。
インドインクの内皮下注入は、これらの線維性シートを観察し、非獣類の心臓における右および左束枝とその突出部の心内膜下コースを示すために用いられる方法の一つである(図6bおよびd)。 ヒツジと子牛の心臓を調べたところ、これらはヒトの心臓とはいくぶん異なることがわかりました。 子牛の心臓は、左束枝の束が通常3本で、心室間中隔の上部に発生する点でヒトの心臓に似ている(図6b)。 しかし、ヒツジの心臓では2本の束しか見られず、しかもそれが中隔壁の長さの中途に現れている。 ヒツジと子牛の心臓では、小さな筋小胞体が心室腔を横断しており、いわゆる「偽腱」と呼ばれるもので、その内部でHis枝の遠位端を乳頭筋と隣接する心室壁に向かって運んでいる。 心臓の右側では、羊と子牛の心臓のモデレータバンドはヒトのものよりも細長いが、その内部には常に右束枝の分枝がある(図6c)。
無蹄類の心臓では、心内膜下プルキンエネットワークは、左心室でも右心室でも楕円状に配列されている(図6e)。 また、その輪郭から心室壁を貫通する枝が生じ、新しい枝や他の枝との吻合に至る(図6e)。 Oosthoekらの解剖学的および免疫組織化学的研究によると、ウシ心臓では、プルキンエ線維が結合組織の被覆を失ったところに、ごく小さな移行細胞の領域が存在することが示されている31。 プルキンエ線維が結合組織の被覆を失うと、電気インパルスはCSから心室の作業筋細胞へと伝わる。 心室壁における作業筋原線維の空間的配向が、心室伝導の異方性を決定する(図6f)。
結論 種によって違いはあるものの、結節の構造、およびヒト房室伝導系の残りの部分は、通常使用する実験動物のものと同様である。 心臓インパルスを発生させる構造であるSA結節は、右心房の一方の端に位置している。 SAノードからのインパルスは、心房壁の後方を、心房の末端部に到達するまで、心房繊維の複雑かつ正確な空間配置を通って伝搬する。 この終点で房室結節の移行細胞がインパルスを受け取り、Hisバンドルを介して伝達される前にそれを遅延させる。 房室結節は心房と心室の間の絶縁線維面を横切り、2つの枝(右房室枝と左房室枝)を経由して対応する心室へインパルスを伝達する。 これらの枝は、それぞれ心室筋細胞の結合鞘によって絶縁されている。 このような配置により、特殊な筋細胞と作業筋細胞との接触は、His束の遠位部においてのみ可能である。 このように、房室伝導系は、約100年前に俵2によって主に記述されたもので、心臓のインパルス伝達に秩序を与える構造になっている。 心臓内の特定の伝導組織の構造と位置を知ることは、さまざまな心調律の乱れの解決に役立つ可能性がある。
Leave a Reply