A szívcsomók és az atrioventricularis specializált ingerületvezetési rendszer anatómiája | Revista Española de Cardiología
INTRODUCTION
Stannius1 1852-es klasszikus tanulmányai voltak az elsők, amelyek azt javasolták, hogy a szív ingerületvezetése myogén. Körülbelül egy évszázaddal ezelőtt kimutatták, hogy a szívverés megindulásáért és terjedéséért specializált izomszövet felelős. 1906-ban Sunao Tawara2 megerősítette a His3 által még 1893-ban leírt izomköteg létezését. Ugyancsak 1906-ban Keith és Flack4 megerősítette a His-Tawara-rendszer létezését. Egy évvel később leírták a sinoatrialis (SA) csomó szerkezetét.5
Bár Purkinje6 írta le elsőként a speciális kamrai rostokat, nem volt tisztában azok jelentőségével a szív szerkezetében, és Tawara2 volt az, aki kimutatta, hogy a His által leírt izomköteg folytonos a kamrai Purkinje-rostokkal.6 Tawara tanulmányait nemrégiben fordították le angolra,7 bár munkája egy részének első fordítását erre a nyelvre Robb8 vállalkozott 1965-ös tankönyvében. Ezek a munkák alapvető olvasmányok minden olyan kutató számára, aki a szív ingerületvezetési rendszerét (CS) tanulmányozná.
A SPECIÁLIS ATRIOVENTRIKULÁRIS VEZETÉSI RENDSZER KOMPONENSEI
A pitvarokat a kamrákkal összekötő izomköteget His3 “átható kötegként” írta le. His azonban nem figyelte meg ennek a kötegnek a szövettani folytatását a jobb pitvarban a pitvari-ventrikuláris (AV) csomóval, a kamrákkal vagy a kamrai Purkinje-sejtekkel. Tawara2 volt az, aki ezt a kapcsolatot felismerte, miközben Aschoff irányításával doktorált. Korábban, 1893-ban Kent9 és His3 is leírták az izmos AV-kapcsolatokat, amelyek sokáig nagy zavart okoztak.10 Később megfigyelték, hogy ezek a kapcsolatok nem az egészséges, hanem a beteg szívekben találhatók meg.
Kent és His megállapításai nyomán a klinikai kardiológusok és a fiziológusok egyaránt keresték a szívimpulzus generálásáért felelős szerkezetet. Azt gyanították, hogy ez a vena cava superior és a jobb pitvar találkozásának területén található; kísérleti körülmények között ez a szívnek az a része, amely utoljára áll le a szívverés (az úgynevezett ultimum moriens). 1907-ben Keith és Flack5 minden általuk vizsgált emlősben, így az emberben is megkülönböztette az SA- vagy szinuszcsomót. Úgy vélték, hogy az őt alkotó sejtek a szívimpulzus kiindulási helye.
A CS az SA-csomóban keletkezik, amely a jobb pitvar felső elülső részén található (1. ábra). Az AV-csomó az alsó, hátsó pitvarban található. A CS az AV-csomópontból a behatoló His-kötegbe nyúlik, majd a bal és a jobb köteg ágakra oszlik, amelyek az interventricularis septumon keresztül ereszkednek le, egy kötőszöveti hüvelybe burkolva, amely elszigeteli őket a környező izomszövettől. A szívizomzaton belül folytonosak a Purkinje-hálózattal (1. ábra).
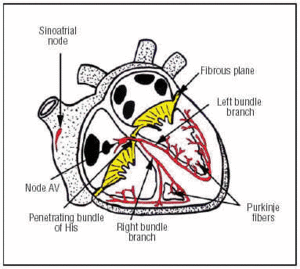
1. ábra. A szív ingerületvezetési rendszer diagramos ábrázolása (piros). A behatoló Bundle of His átfúrja a rostos atrioventricularis (AV) síkot.
Morfológiai-makroszkópos érdekességek
Több makroszkópos érdekesség segít a szív CS helyének meghatározásában. Az SA-csomó, amely szubepikardiális (2a. ábra), a vena cava superior és a pitvari függelék izomzata közötti csomópontba ékelődik. Alapja a terminális gerinccel szemben helyezkedik el. Az SA-csomó és az epikardium közötti távolság 0,3±0,1 mm.11 A személyek körülbelül 10%-ánál a csomó nem terjed a vena cava inferior felé, hanem patkó alakban fekszik a vena cava superior nyílásának alsó része körül.12 Az AV-csomó a pitvari septum alján, egy Koch által először bemutatott háromszög alakú terület csúcsán található.13 Ez a háromszög a jobb pitvar endocardialis felszínén helyezkedik el (2b. és c. ábra), elölről a tricuspidalis billentyű szeptumlapjának behelyezése, hátulról pedig egy Todaro-ínként ismert rostos ín határolja. Ez az ín az Eustachian billentyű rostos subendocardialis folytatása, és a pitvari izomzatba illeszkedik, amely elválasztja a sinus coronaria nyílását a fossa ovale-tól. E háromszög csúcsát felfelé a fent említett elülső és hátsó határ találkozása alkotja, ami megfelel a szív központi rostos testének (CFB). A háromszög alapját a sinus coronaria nyílása és a jobb pitvarnak a tricuspidalis billentyű szeptális lapocskáját alátámasztó előcsarnoka alkotja. Ezt az alapot az elektrofiziológusok szeptális isztmusként ismerik, és itt végzik a lassú pálya rádiófrekvenciás ablációját AV-csomós reentráns tachycardia esetén.14
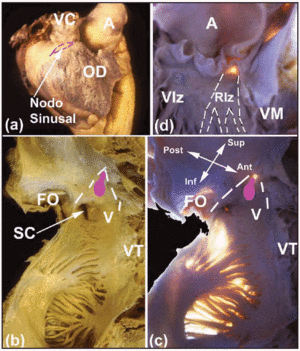
2. ábra. A: A jobb pitvar laterális epikardiális nézete, az SA-csomó helyét a szaggatott rózsaszín vonal mutatja. B és C: a jobb pitvar hátsó és szeptális falának endokardiális nézetei (normál és transzilluminációval), amelyeken látható az ovális fossa (OF) és a Koch-háromszög határai (szaggatott fehér vonalak), a Todazo-ín (TT) és a tricuspidalis billentyű (TV) szeptális csúcsának behelyezése. A jobb pitvar előcsarnoka (V) és a koszorúér-szinusz (CS) nyílása alkotja az alsó határt. Az AV-csomó elhelyezkedését egy ovális csomó (rózsaszín) mutatja. D: bal kamrai nézet a hártyás septum bemutatására (transzillumináció). Ez a His-köteg kilépési pontja és folytonossága a jobb és bal köteg ágakkal. A bal kötegág (LBB) szaggatott fehér vonalakkal van jelölve. A jelöli az aortát; AA, pitvari függelék; VC, felső vena cava; RV, jobb kamra; LV, bal kamra; MV, mitrális billentyű.
Az AV-vezetés folytatása a behatoló His-kötegen keresztül történik, amely a vezetési tengely egyetlen olyan része, amely a CFB-t perforálja. A CFB-t az aorta- és a mitrális szívbillentyűszárny kötőszövetének a tricuspidalis billentyű szeptumszárnyával – az úgynevezett jobb rostos trigon – és az interventricularis septum hártyás részével való egyesülése alkotja. Sok emlősszívben a trigonus rostos, de a szarvasmarhaszívnek van egy központi csont- vagy porctömege (os cordis). Ezzel szemben az ámbráscet CFB rostos szövete nagyon laza. A hártyaszerű rész vagy septum, amely különböző hosszúságú lehet, jó útmutató a His AV-köteg helyének meghatározásához. Ez e hártyás rész felett jelenik meg a jobb oldali rostos trigonon való áthaladás után (2d. ábra), majd a bal és a jobb köteg ágaira oszlik. A jobb ág a jobb kamra medialis papilláris izomzatának tövében a szeptális izomzaton keresztül halad át. Ezután vékony vezetékké válik, amely mélyen behatol a septomarginalis trabeculatio vagy a medialis és az elülső papilláris izmokat összekötő mérsékelt sávba. A bal ág eredése az aortabillentyű jobb és nem-koszorúér-csúcsok közötti commissura alatt található; ezután az interventricularis septum subendocardiumán keresztül ereszkedik le (2d. ábra). Útja néha látható az azt burkoló fényes rostos lamina miatt. A bal ág proximális része sokkal hosszabb, mint a jobb ágé. Alkalmanként egy harmadik, “holtágnak “15 nevezett ág is megfigyelhető a magzati vagy csecsemőszívekben, amely a His-köteget anterio-superior irányban folytatja az aorta gyökere felé.
A csomópontok és az atrioventrikuláris ingerületvezetési rendszer felépítése
Az olyan vizsgálatok, amelyekben a szövettani technikák hasonlóak voltak a Tawara2 és a későbbi munkások, mint Davies16 és Truex et al17 (hogy csak néhányat említsünk) által alkalmazott technikákhoz, kimutatták, hogy az ember CS-je egészen hasonlóan épül fel, mint más emlősöké (kisebb eltérésekkel a fajok és a szívek között). Tawara2 arról számolt be, hogy a specializált szívizomsejteket egy vékony, fénymikroszkóp alatt látható kötőszöveti lemez választja el a normál vagy működő szívizomsejtektől, és ezen alapulnak az Aschoff18 és Mönckeberg19 által javasolt kritériumok a specializált szívizom szövettani azonosítására. Egyszerűen fogalmazva, a specializált myocyták fénymikroszkóp alatt vizsgálva elkülönülnek a dolgozó myocytáktól, és szövettani metszetről szövettani metszetre “követhetők”. Monográfiájában Robb8 a vezetőszövetet inkább az “összekötő”, mint a “vezető” rendszer kifejezéssel határozta meg, mivel a szövettani preparátumok jobban meghatározzák a sejtmorfológiát, mint a funkciót. Ő is különbségeket figyelt meg a specializált szívizomzat textúrájában a boncolási anyag frissességétől és az alkalmazott rögzítési és festési módszerektől függően. Tawara2 tisztában volt ezzel, és rámutatott a specializált szívizomsejtek morfológiájának heterogenitására még a szív szövettani metszeteiben is. Egy adott fajon belül a legszembetűnőbb különbségek a vizsgált egyedek életkorával függnek össze.20 Az utóbbi években különböző molekuláris és immunhisztokémiai markereket használtak az ember és más emlősök embrionális szívében a vezetőszövet lokalizálására. Nem találtak azonban olyan specifikus markert, amely ezt a szövetet felnőtt emberben kiemelné.
A normális emberi szívben az SA- és AV-csomók nem felelnek meg az Aschoff és Mönckeberg18,19 kritériumainak, mivel nem szigeteli őket kötő- vagy zsírszövet elektromosan a környező szívizomtól. Inkább egy kis, átmeneti sejtekből álló terület után érintkeznek a pitvari munkarostokkal. Az SA-csomópontban Keith és Flack5 különbséget tett a szinusz- és a munkasejtek között. Tawara2 azonban jelezte, hogy milyen nehézségekbe ütközött az AV-csomó sejtjeinek megkülönböztetése a His köteg sejtjeitől. Ezért azt javasolta, hogy a köztük lévő különbség pusztán anatómiai jellegű. E meghatározás alapján a CS-nek a CFB által teljesen burkolt részét nevezzük átható kötegnek vagy His-kötegnek (3a. ábra). A proximális ingerületvezetési rendszertől a His kötegig terjedő pitvari részt AV-csomónak nevezik (3b. ábra). Ez az anatómiai megkülönböztetés logikus, mivel a behatoló His-köteg szigetelése megakadályozza, hogy közvetlen kapcsolatba kerüljön az afferens pitvar elektromos aktivitásával. Ezért minden pitvari aktivitást előzetesen az AV-csomón keresztül kell irányítani.
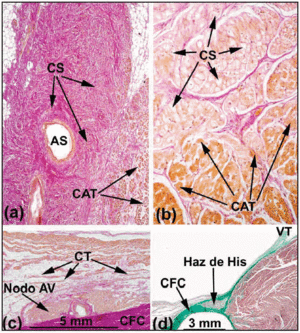
3. ábra. Emberi (a;x10) és sertésszív (b;x40) sinoatrialis (SA) csomópontjának van Gieson-módszerrel festett sagittalis szövettani metszetei. Figyeljük meg a szinuszsejtek (SC) és a dolgozó pitvari sejtek (WAC) közötti kapcsolatot. A sinus sejteket az jellemzi, hogy világosabbak és nagyobb mennyiségű kötőszövetbe ágyazottak (piros). c: van Gieson-festésű metszet a Koch-háromszög középső zónájából. Figyeljük meg a kompakt AV-csomópont alakját és a kompakt csomópont domború felszínével érintkező átmeneti sejteket (TC). d: Masson trikróm-festésű metszet, amely a CFB rostos szövetekkel (zöld) körülvett, behatoló His-kötegét mutatja. Az SNA a sinuscsomó artériát jelöli; CFB, központi rostos test; TV, tricuspidalis billentyű.
A SA-csomó eredendő funkciója, hogy a szívimpulzus forrása legyen. Az SA-csomópont az emberben íves vagy fusiform struktúra. Histológiailag a normál működő sejteknél valamivel kisebb sejtekből áll, amelyek kötegekbe rendeződnek. Ezek térbeli rendezettség nélkül keverednek egymással, gyengén festődnek, és sűrű kötőszöveti mátrixba ágyazódnak (3. a és b ábra). Az életkor előrehaladtával a kötőszövet mennyisége a csomósejtek által elfoglalt területhez képest növekszik.21 A csomó perifériáján a specializált sejtek keverednek a dolgozó szívizomzat sejtjeivel (3a. és b. ábra). Ezenkívül többszörös, a dolgozó pitvari szívizommal interdigitáló sugarakat vagy nyúlványokat írtak le. Ezek intramyocardialisan behatolnak a terminális gerincbe, valamint a vena cavae superiorba és inferiorba. Az SA-csomó egy artéria, az úgynevezett szinuszcsomó-artéria körül helyezkedik el, amely a csomóponton belül centrálisan vagy excentrikusan futhat. Az emberi szívek 29%-ában ez az artéria a csomóponton belül ágazik el.11 Az SA-csomópont szoros kapcsolatban áll a vegetatív idegrendszerrel is. Azt feltételezik, hogy ezen idegrostok többsége paraszimpatikus, a szimpatikus rostok a csomóponti erek körül koncentrálódnak.23
Az AV-csomópont eredendő funkciója a szívimpulzus késleltetése. Emberben ez a csomópont egy kompakt részből és egy átmeneti sejtekből álló területből áll. Az előbbi félig ovális és a CFB felett fekszik (3c. ábra). A Koch-háromszög alapjához közeli szakaszokon a csomópont kompakt része két nyúlványra vagy nyúlványra oszlik. Az AV-csomót vaszkularizáló artéria általában ezek között található. E nyúlványok hossza szívenként változó.24 Az átmeneti sejtek mérete köztes az AV-csomó és a pitvari munkasejtek között. Nagyobb mennyiségű kötőszöveti sejt veszi őket körül, mint ami a munkasejteket borítja, de nincsenek elszigetelve a szomszédos szívizomtól. Inkább egyfajta hidat képeznek a dolgozó és a csomóponti szívizom között, és összegyűjtik az elektromos információt a pitvari falakról, és továbbítják azt az AV-csomóba.
Vitatott, hogy az SA-csomóból érkező impulzus hogyan jut el az AV-csomóba. Egyes szerzők a köztük lévő speciális pályák létezését feltételezik.25 Vizsgálataink nem támasztják alá ezt az elképzelést, hanem azt a hipotézist támogatják, hogy maguk a dolgozó izomrostok (és azok geometriai elrendeződése a pitvarfalakban) felelősek azért, hogy a vezetés a pitvar egyes területein gyorsabb, mint máshol.26
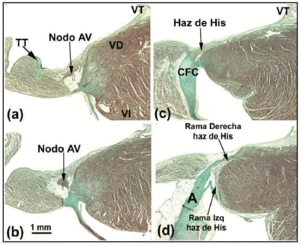
Az AV-csomópont distalisan a behatoló His-köteggel folytatódik (3d. ábra), bár e két struktúra között a sejtek elrendeződését tekintve vannak kisebb különbségek, beleértve a His-köteg sejtjeinek párhuzamosabb módon való elhelyezkedését. Ennek magyarázata morfológiai lehet: a His-köteget a CFB kötőszövete kezdi körülvenni, így válik vezetőképes traktussá, amely információt visz a kamrákba.
A kutya AV-csomója kisebb, mint az emberé, de hosszabb átható His-köteggel rendelkezik.27 Egyes szerzők28 ezt úgy értelmezik, hogy a kutya AV-csomójának egy része a CFB-n belül helyezkedik el. A nyúlnál más szerzők29 a His köteg egy részét úgy írják le, mintha az az AV-csomó részét képezné, de ez tévedés (4a-d. ábra). A kutya, a nyúl és az ember AV-csomója között a legkiemelkedőbb morfológiai különbség az, hogy az előbbit nem borítják átmeneti sejtek. Patkányokban (amelyek nyugalmi szívfrekvenciája 10-szer gyorsabb, mint a kutyáé vagy az emberé) az AV-csomó arányaiban hasonló a kutyáéhoz, de a CFB kisebb.
4. ábra. Ez az összetett ábra a nyúlban az atrioventricularis (AV) csomót, valamint a His köteget és annak jobb és bal kötegágát mutatja. A b-ben a vízszintes sáv 1 mm-t jelöl (minden képen azonos). Masson trikróm festés. A az aortát jelöli; TT, Todaro ín; RV, jobb kamra; LV, bal kamra; TV, tricuspidalis billentyű.
Ha a vezetési rendszer szövettani pályáját követjük a behatoló His-köteg felé, sok emberi szívben látható, hogy ez utóbbi balra fordul, és az intertricularis septum izmos gerincén lép ki. A CFB kötőszövetével körülvéve a His-köteg hossza változhat, mielőtt a bal és a jobb köteg ágra szétválna. Az előbbi ág az interventricularis septum bal oldalán halad át (5a. és c. ábra). A His-köteg osztódása egy zsokéra hasonlít, amely az interventricularis septum izmos gerince fölött guggol (5a. ábra). Alkalmanként azonban balra deviál (5c. ábra). Ilyenkor a jobb ág a septum izomzatának belsejébe lép (5b. ábra), és a jobb kamrában a medialis papilláris izom beékelődésével együtt jelenik meg.
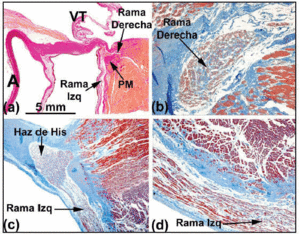
5. ábra a: van Gieson-festésű metszet, amely a bundle of His-t (emberi szív) az interventrikuláris septum hártyás része (MP) felett mutatja. b: Eredeténél a jobb ág ebben a szívben intramyocardialis és kötőszövet (kék) veszi körül (Jones trikrómfestés, x20). c: a His-köteg osztódása ebben a szívben az interventricularis septum izmos gerincének bal oldala fölé tolódik, és hosszanti irányban a bal kamra endocardiuma alá ereszkedik (Jones´ trikrómfestés, x5). d: figyeljük meg a bal ág fedő kötőszövetes hüvelyét (kék) (Jones´ trikrómfestés, x10). A jelöli az aortát; E, endokardium; TV, tricuspidalis billentyű.
A jobb és bal köteges ágakat proximális szakaszuk mentén rostos lamina fedi (5b és d ábra). Amint Tawara2 kimutatta (6a. ábra), emberben a bal ág jellemzően három fascikulára oszlik, amelyek között kiterjedt összeköttetés van. Ezek a fasciculák a kamracsúcsban elágaznak, és a mitrális billentyű két papilláris izmának belsejébe, de a kamrafalak mentén a szívalap felé is visszahúzódnak. Távolabb, az emberi szív kamráinak csúcsában szinte lehetetlenné válik a Purkinje-rostok elágazásainak nyomon követése, mivel ezek elveszítik rostos bevonatukat, és nagyon hasonlítanak a működő szívizomzatra.
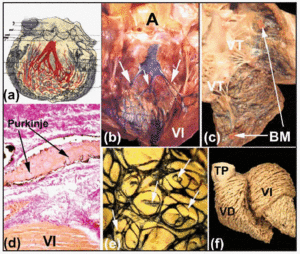
6. ábra a: Tawara ábrája, amely a bal kötegág trifaszkuláris elrendeződését mutatja az emberben. b és c: friss borjúszív, amelyben a jobb és bal kamrát megnyitották. A tinta szubendocardialis injekciói feltárják a jobb és bal köteges ágakat és a Purkinje-hálózatot. Figyeljük meg B-ben a bal köteg ág három fascikuláját (nyilak), C-ben pedig a moderátor sávot (MB). d: egy borjúszív metszete (van Gieson festés, x100), amelyet a Purkinje-hálózatba adott tintainjekció után kaptunk), amelyet az eredésénél kötőszövet (piros) burkol be. e: a Purkinje-hálózat szubendocardialis elrendeződése egy borjú bal kamrájában. Figyeljük meg a hálózat elliptikus elrendeződését és a széleiről kiinduló, a szívizomzatba behatoló mellékágakat (nyilak). f: felnőtt emberi szív kamrai izomrostjainak boncolása. Figyeljük meg a bal kamra mediális és mély rétegei közötti elrendeződési különbséget. A jelöli az aortát; PT, tüdőtörzs; LV, bal kamra; LV, bal kamra; TV, tricuspidalis billentyű.
A tinta szubendocardialis injekciója az egyik olyan módszer, amelyet e rostos lapok megfigyelésére és a jobb és bal kötegágak szubendocardialis lefolyásának és elágazásainak bemutatására alkalmaznak patás szívekben (6b. és d. ábra). A juh- és borjúszíveken végzett vizsgálataink azt mutatják, hogy ezek némileg eltérnek az emberi szívektől. A borjúszívek jobban hasonlítanak az emberi szívhez abban, hogy a bal köteg ág fasciculái általában három darabból állnak, és az interventricularis septum felső részén erednek (6b. ábra). A juhszíveknél azonban csak két fascikula található, és ezek a szeptumfal hosszának felénél jelennek meg. Mind a juh-, mind a borjúszívben kis izmos trabeculák keresztezik a kamrai üreget – az úgynevezett “hamis ín” -, amelyek belsejükben a His-ágak disztális elágazásait a papilláris izmok és a szomszédos kamrafalak felé viszik. A szív jobb oldalán mind a juh-, mind a borjúszív moderátorszalagja karcsúbb, mint az emberé, de belül mindig a jobb köteg ág egy-egy mellékágát tartalmazza (6c. ábra).
A patások szívében a szubendocardialis Purkinje-hálózat elliptikus elrendezésű, mind a bal, mind a jobb kamrában (6e. ábra). Ezenkívül a kontúrjából olyan ágak erednek, amelyek áthatolnak a kamrafalon, és új ágakhoz vagy más ágakkal való anasztamózisokhoz vezetnek (6e. ábra). A Purkinje-hálózat intramurális elágazásait azonban emberi szívben nem mutatták ki.30
A Purkinje-hálózattal kapcsolatos ellentmondásos pont a működő kamrai szívizomzat és a Purkinje-rostok közötti átmeneti sejtek létezése.31 Oosthoek és munkatársai anatómiai és immunhisztokémiai vizsgálatai30 azt mutatják, hogy szarvasmarha szívben az átmeneti sejtek egy nagyon kis zónája van, ahol a Purkinje-rostok elveszítik kötőszöveti borításukat. Ilyen sejteket azonban juhszívben nem figyeltek meg.32 Amikor a Purkinje-rostok elveszítik kötőszöveti borításukat, az elektromos impulzusok a CS-ből a kamrák működő myocitáihoz jutnak. A dolgozó myofibrillumok térbeli orientációja a kamrafalakban határozza meg a kamrai ingerületvezetés anizotróp jellegét (6f. ábra).
KÖVETKEZTETÉSEK
Bár fajok között vannak különbségek, a csomópontok, valamint az emberi AV-vezetési rendszer többi részének felépítése hasonló az általánosan használt laboratóriumi állatokéhoz. Az SA-csomópont, a szívimpulzust generáló struktúra, a jobb pitvar egyik végpontján helyezkedik el. A belőle kiinduló impulzusok a pitvarfalakban hátrafelé haladnak a dolgozó pitvari rostok bonyolult, de pontos térbeli elrendezésén keresztül, amíg el nem érik a pitvar végét. Ezen a végén az AV-csomó átmeneti sejtjei fogadják az impulzust, és késleltetik azt, mielőtt az továbbításra kerülne a His-kötegen keresztül. Ez utóbbi keresztezi a pitvarok és kamrák közötti szigetelő rostos síkot, és két ágon (a jobb és bal kötegágon) keresztül továbbítja az impulzust a megfelelő kamrák felé. Mindegyik ágat a dolgozó kamrai szívizomsejtek kötőhüvelye szigeteli. Ez az elrendezés a specializált és a dolgozó szívizomsejtek között csak a köteg distalis elágazásainál teszi lehetővé az érintkezést. Ily módon a Tawara2 által közel 100 évvel ezelőtt nagyrészt leírt AV-vezetési rendszer úgy van felépítve, hogy rendet teremtsen a szívimpulzusok továbbításában. A szívben lévő specifikus vezetőszövetek szerkezetének és elhelyezkedésének ismerete segíthet megoldást nyújtani a szívritmus különböző zavaraira.
Leave a Reply