América del SurPerro de monte Speothos venaticus

perro de monte – © Gerald L Zuercher
Grupo de Trabajo de Cánidos Amazónicos – Karen DeMatteo y Fernanda Michalski son las coordinadoras del Grupo de Trabajo de Cánidos Amazónicos. Este grupo de trabajo se centra en cuatro especies: el perro de orejas cortas, el zorro cangrejero, el perro de monte y los zorros sudamericanos. Los cánidos amazónicos son similares a muchos carnívoros de todo el mundo cuya supervivencia a largo plazo se ve amenazada por una serie de amenazas directas e indirectas, como la pérdida de hábitat, la expansión de las presas hidroeléctricas, la caza ilegal de presas y las enfermedades de los perros domésticos.
Proyectos
- Proyecto Zorro Pitoco: Utilización de técnicas no invasivas para aportar nuevos conocimientos sobre los perros de monte y la biodiversidad coexistente en Misiones, Argentina
- Monitoreo de la fauna con cámaras trampa en el Parque Nacional Madidi y sus alrededores, Bolivia
- Investigación de helmintiasis neotropicales y otros parásitos en el biotopo de animales silvestres de la Selva Misionera, como potenciales focos de zoonosis
- Efecto del cambio climático en los carnívoros de la Amazonía
Enlaces relevantes
- Flickr: Cánidos salvajes del mundo
- Evaluación de la Lista Roja de la UICN 2011 – Perro de monte
Informes / Documentos
- Residentes de la selva – cánidos del Amazonas
- Encuesta de estado 2004 &Plan de acción de convervación -. América del Sur
- Por qué los perros de monte son tan diferentes de otros perros
- Raros perros de monte del Amazonas captados por una cámara
- Un perro de rescate ayuda ahora a salvar a otros perros (mucho más salvajes)
- Especies sorprendentes: perro de monte
Otros nombres
En inglés: Bush Dog, Savannah Dog, Vinegar Dog
Francés: Chien Des Buissons, Zorro
Español; castellano:Cachorro Vinagre, Guanfando, Pero Selvático, Perrito Venadero, Perrito de Monte , Perro De Monte, Perro Grullero , Perro Vinagre , Perro de Agua, Perro de la Selva, Umba, Zorrito Vinagre, Zorro Pitoco , Zorro Vinagre
Alemán: Waldhund
Italiano: Itticione, Speoto
Portugués: Cachorrinho, Cachorro-do-mata, Cachorro-do-mato-cotó, Cachorro-do-mato-vinagre, Cachorro-pitoco
Justificación
Aunque extendido, el perro de monte parece ser raro en toda su área de distribución. La especie ha demostrado ser extremadamente difícil de localizar en la naturaleza, lo que dificulta la estimación de las tendencias de la población. Aunque se podría esperar que una especie ampliamente distribuida con apariciones confirmadas en hábitats fragmentados o alterados tuviera una población generalmente estable, las pruebas preliminares obtenidas sobre el terreno sugieren que estas áreas pueden alterar los requisitos ecológicos del Perro de los Bosques, lo que resulta en un aumento del área mínima necesaria para mantener un grupo (y, por lo tanto, pone a la especie en mayor riesgo de exposición directa o indirecta a otras amenazas). La especie se clasifica aquí como Casi Amenazada basándose en una disminución estimada del 20-25% en los últimos 12 años (duración estimada de la generación = 4 años), aproximándose así al umbral para su inclusión en la lista de Vulnerables según el criterio A2. Este declive es atribuible a múltiples amenazas, incluyendo la pérdida y fragmentación del hábitat, la reducción de la abundancia de presas debido a la caza ilegal y la depredación por parte de los perros domésticos, y un mayor riesgo de enfermedades relacionadas con los cánidos.
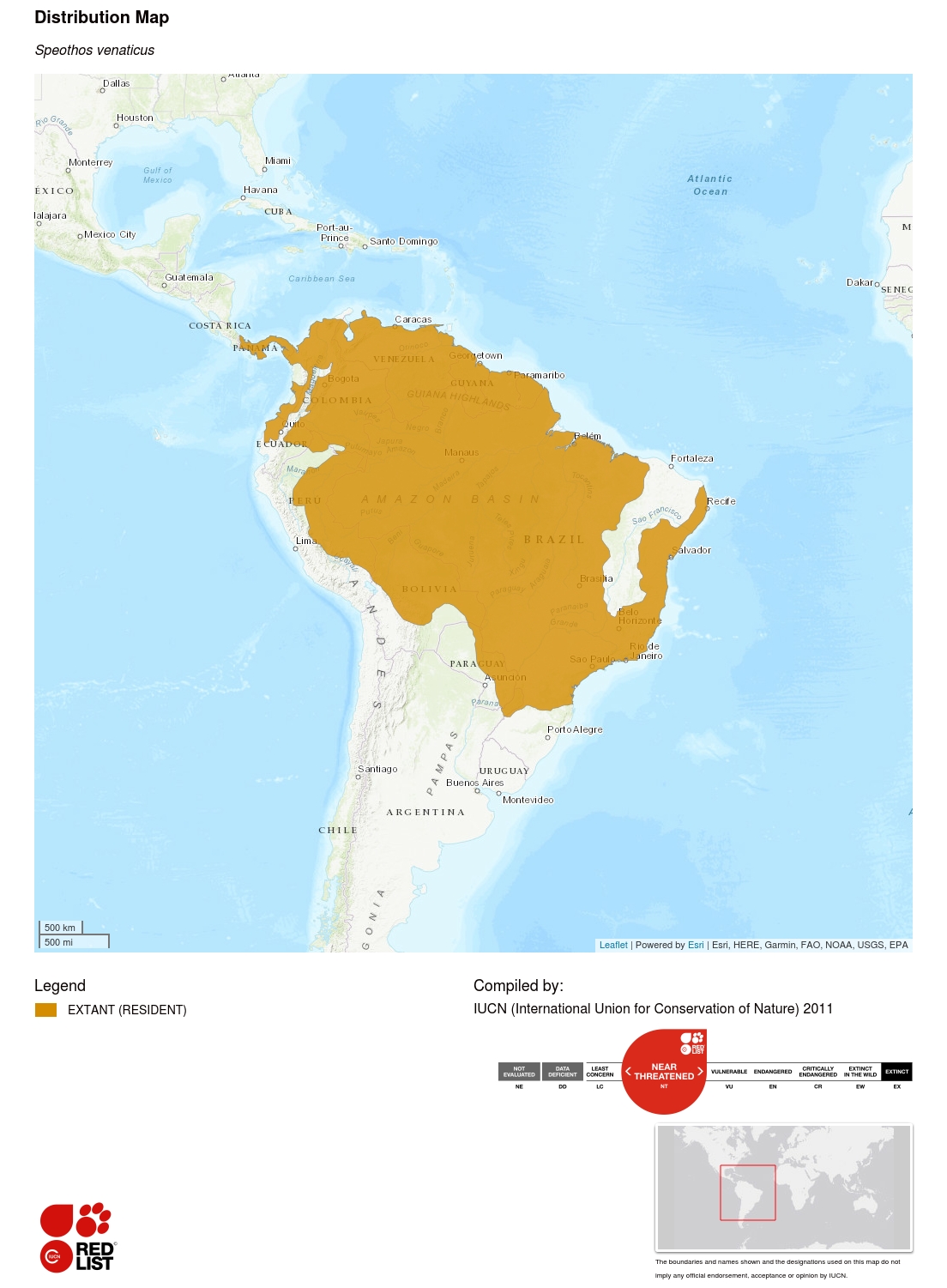
Información sobre el área de distribución geográfica
Esta especie se encuentra desde el extremo oriental de América Central y el norte de América del Sur, al sur de Paraguay y el noreste de Argentina. Es posible que aún existan subpoblaciones aisladas en Ecuador y Colombia, al oeste de los Andes. Su distribución histórica puede haberse extendido hasta el norte de Costa Rica (Rosa y Nocke 2002), donde la especie aún puede sobrevivir. El mapa de distribución actual se generó a partir de los resultados de un amplio estudio de biólogos de carnívoros y de la literatura sobre la presencia de la especie (n = 399 localidades históricas), así como de un modelo bioclimático que predice el área adecuada para la especie (DeMatteo y Loiselle 2008). Estos últimos autores estimaron que el área total que se predice como adecuada para los perros de monte es del orden de 14.445.000 km², de los cuales el 77% se encuentra dentro del área de distribución actual.
Tendencia de la población:Disminución
Información sobre la población
A pesar de su amplia área de distribución y de su presencia en una gran variedad de hábitats, los perros de monte parecen ser naturalmente raros en toda su área de distribución. En una encuesta reciente realizada por DeMatteo (2008), la mayoría de los países en los que se distribuye el Perro de monte informaron del estado de la especie como rara o desconocida (rara: Ecuador; rara o desconocida: Argentina, Bolivia, Brasil, Guayana Francesa, Paraguay; desconocido: Panamá y Venezuela) y sólo dos países la reportaron como común (Guyana y Perú). La especie rara vez se registra con cámaras trampa; sin embargo, hay que tener cuidado al interpretar la falta de detección, ya que esto puede estar asociado a que la especie evita activamente las características asociadas a esta técnica (por ejemplo la colocación de las trampas a lo largo de los senderos de los animales utilizados por otros carnívoros) o las suposiciones sobre el movimiento de los animales (por ejemplo, la colocación a lo largo de las carreteras asume que el animal caminará a lo largo de la carretera en lugar de cruzar por encima de la carretera).
Los datos demográficos para los perros de monte son escasos en toda su área de distribución y las estimaciones de la población se han reportado sólo para unas pocas áreas: <100 en Misiones Argentina (DeMatteo 2008), >1.000 en Bolivia (DeMatteo 2008), >1.000 en 4.022 km² o un individuo/4 km² en Cusco Perú dentro de la región del río Camisea (DeMatteo 2008), 0,04 individuos/km² (B. Beisiegel com. pers.) en Brasil, 0,025 individuos/km² en un cerrado parcialmente fragmentado en Mato Grosso Brasil (E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima y S. Klorfine, obs. pers.), 15,8 individuos/394 km² o 0,04 individuos/km² en el Pantanal en Brasil (Lima, Jorge et al. 2009), y 0,001 individuos/km² en la fragmentada Amazonia meridional de Brasil (Michalski 2010). Un área adecuada de aproximadamente 11.000.000km² (DeMatteo y Loiselle 2008), y una densidad de población media de aproximadamente 0,01 individuos/km², predecirían una población estimada de 110.000 individuos, de los cuales aproximadamente la mitad serían maduros; sin embargo, los efectos de la fragmentación, el grado de protección y la variabilidad a lo largo de su amplia distribución sugerirían que esto es una sobreestimación.
En toda la distribución del Perro de monte, las tendencias de la población se reportan como desconocidas en dos países (Guyana y Panamá), en disminución en dos (Paraguay y Venezuela), desconocidas o estables en uno (Guayana Francesa), y estables en dos (Ecuador y Perú, específicamente Cusco Perú) (DeMatteo 2008). En toda el área de distribución, los crecientes niveles de fragmentación del hábitat (urbanización y agricultura), las amenazas a las poblaciones de presas con la caza furtiva ilegal y la exposición a enfermedades potencialmente letales relacionadas con el cándido, hacen que la probabilidad de que la tendencia se mantenga estable o aumente sea altamente improbable.
Información sobre el hábitat y la ecología
Los perros de monte son reportados como generalistas de hábitat por los pueblos indígenas, dentro del contexto de ocurrir generalmente cerca de las fuentes de agua, particularmente pequeños arroyos, donde las densidades de presas pueden ser más altas (Zuercher et al. 2005; E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima y S. Klorfine obs. pers.; M. Swarner, com. pers.). Los perros de monte han sido observados en hábitats boscosos de tierras bajas (por debajo de 1.500 m), incluyendo bosques primarios y de galería (Defler 1986), bosques semicaducifolios y bosques estacionalmente inundados (Aquino y Puertas 1997). También se han registrado observaciones en el hábitat del cerrado en Brasil (Silveira et al. 1998; Oliveira 2009, C. Brady com. pers.) y en Paraguay (Zuercher y Villalba 2002) y en áreas de borde de la pampa (sabana húmeda) y ribereñas (Strahl et al. 1992, Emmons 1998). Informes recientes mencionan a los perros de monte en tres hábitats únicos: la caatinga, el chaco y los manglares de la costa (DeMatteo y Loiselle 2008). En algunos casos, se han observado a varios kilómetros del hábitat forestal (Silveira et al. 1998). La especie también se reporta ocasionalmente en bosques secundarios, ranchos (M. Swarner com. pers.), ranchos cerrados fragmentados (E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima y S. Klorfine obs. pers.; L. Silveira y A. Jácomo com. pers.), áreas perturbadas (DeMatteo y Loiselle 2008, Oliveira 2009), y áreas forestales fragmentadas (Michalski y Peres 2005, Michalski 2010).
No se encontró preferencia de selección de hábitat ni en el Pantanal en Brasil (Lima, Jorge et al. 2009) o en la mezcla de bosque atlántico y cerrado del Alto Paraná en Paraguay (Zuercher et al. 2005); sin embargo, se han encontrado algunas evidencias que sugieren una preferencia por el hábitat de sabana y bosque intactos frente a los alterados (tierras de cultivo y pastos) para un grupo de perros de monte en el cerrado parcialmente fragmentado, lo que indica que el uso y la preferencia del hábitat pueden diferir según la región y la disponibilidad (E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima, and S. Klorfine pers. obs.). Mientras que Michalski (2010) reportó una ausencia de Perros de monte en áreas fragmentadas en el sur de la Amazonia, otros estudios han demostrado una fuerte evidencia de que su ocurrencia es probable (DeMatteo y Loiselle 2008; Oliveira 2009; E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima, y S. Klorfine obs. pers.); sin embargo, se desconoce la intensidad de uso en estas áreas y el impacto en los requerimientos ecológicos. Los datos preliminares de campo sugieren que a medida que el grado de fragmentación del hábitat aumenta también lo hace el área requerida por los perros de monte (E.S. Lima com. pers.); aunque estos datos sólo están disponibles para el ecosistema del cerrado en Brasil, se sospecha que es cierto para todos los tipos de hábitat y un reflejo de una relación negativa entre la densidad de presas y la fragmentación del hábitat. Un reanálisis de la distribución histórica del Perro de monte (n > 250 localidades) determinó que aproximadamente una cuarta parte (20%) estaba asociada a un hábitat fragmentado/alterado y el porcentaje de hábitat modificado aumentó cuando la estimación se amplió desde el área necesaria para mantener a un solo individuo hasta el área de un grupo de cría (DeMatteo y Loiselle 2008).
La variación reportada en las estimaciones del tamaño del área de distribución está probablemente asociada con la integridad del hábitat y la densidad de presas: 150 km² (Bosque Atlántico del Alto Paraná; Beisiegel 1999), ca. 16 km² (Beisiegel y Ades 2004), y 140 km² (Fixed Kernel 95%) para un grupo de perros de monte (n = 2-4 individuos) en el cerrado (E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima, y S. Klorfine, obs. pers.).
Los datos preliminares de campo apoyan los estudios en cautividad de que los perros de monte existen en grupos familiares con crías de una o más camadas (DeMatteo 2008, Michalski 2010, E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima y S. Klorfine, obs. pers.); sin embargo, se observan regularmente individuos solitarios en diversos tipos de hábitat. La reproducción en la naturaleza es estacional (DeMatteo 2008). La especie parece tener un patrón de movimiento seminómada frente a un verdadero territorio como muchos carnívoros (DeMatteo 2008, E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima y S. Klorfine, obs. pers.) y se ha registrado que vive en simpatría con Atelocynus microtis en Perú y Brasil (Leite Pitman, Beck et al. 2003, Michalski 2010). La duración estimada de la generación es de cuatro años [(0,5 x duración reproductiva de seis años) + madurez sexual al año (en cautiverio; Porton et al. 1987).
Los datos preliminares de campo apoyan los informes anecdóticos de la dieta y la idea de que los perros de monte son altamente carnívoros (Deutsch 1984, Peres 1991); sin embargo, parece haber evidencia de la preferencia de presas basada en la región geográfica y el tipo de hábitat, que probablemente se ve agravada por la variabilidad estacional. Para el área del Pantanal en Brasil, el principal alimento fue el armadillo de nueve bandas (Dasypus novemcinctus; 94%) con pequeños mamíferos, Agoutis (Dasyprocta azarae), y aves que ocurren en menores cantidades; sin embargo, no se registró ninguna fruta (n = 17, identificada por el olor y las huellas; Lima, Jorge et al. 2009). En el bosque atlántico interior de Paraguay, la mayor parte de la dieta estaba compuesta por Agoutis y Paca (Cuniculus paca) con pequeños mamíferos, roedores, reptiles, invertebrados y frutos de Cecropia en menor cantidad (n = 11, identificados mediante ADN mitocondrial; Zuercher et al. 2005). En un centro de reproducción con éxito en el sureste de Brasil, donde se dispone de carne y fruta (plátano y papaya), los animales comen fruta y carne en igual proporción (L. Saboia, com. pers.). Se ha reportado que la depredación de aves de corral ocurre ocasionalmente (DeMatteo 2008).
Información sobre amenazas
Hay varias amenazas graves percibidas, incluyendo: 1) la invasión humana y la pérdida del hábitat intacto debido a la agricultura a gran escala (por ejemplo, la soja), la conversión de la tierra en pastos y las plantaciones a gran escala de monocultivos de árboles (por ejemplo, eucalipto, pino); 2) reducción de la abundancia de presas debido a la caza furtiva ilegal y a la depredación de perros domésticos; y 3) aumento del riesgo de contraer enfermedades letales de los perros domésticos (proximidad a las poblaciones humanas y a los perros de caza (DeMatteo 2008).
Las enfermedades relacionadas con los cánidos son una amenaza no identificada previamente para los perros de monte. Sin embargo, la evidencia de campo apoya que los patógenos pueden ser transmitidos por los perros domésticos, y los efectos pueden ser potencialmente devastadores, principalmente debido a la vida en grupo de la especie (Mann et al. 1980, Steinel et al. 2001, Leite Pitman, Nieto et al. 2003, Jorge, Morato et al. 2007, Jorge, Nunes, et al. 2007, DeMatteo 2008, Jorge et al. 2008, E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima y S. Klorfine, obs. pers.). E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima y S. Klorfine (obs. pers.) observaron este efecto cuando una pérdida de pelo generalizada, que se sospecha que es un tipo de sarna, se extendió gradualmente a través de un grupo de perros salvajes de monte, acabando con todos los individuos. Además de la sarna, el parvovirus y la rabia son enfermedades que afectan negativamente a las poblaciones salvajes (Mann et al. 1980, DeMatteo 2008). Se ha identificado que la especie es susceptible tanto a Dioctophyma renale como a Amphimerus interruptus (colecciones de museo – Vierira et al. 2008), Lagochilascaris sp. (Volcán y Medrano 1991), y como huésped de Echinococcus vogeli (Cestoda: Taeniidae) (Rausch y Bernstein 1972). Se ha identificado que los animales en cautividad son susceptibles a diversas enfermedades y parásitos, como el parvovirus (Janssen et al. 1982), el moquillo canino inducido por vacunas (McInnes et al. 1992), la leishmania (Lima, Fattori et al. 2009), Spirocerca lupi (Rinas et al. 2009), Toxoplasma gondii (Sedlak y Bartova 2006) y Campylobacter (L. Saboia com. pers.). Con este último, 13 de los 15 individuos de un gran grupo familiar murieron repentinamente de una grave enteritis hemorrágica al contraer Campylobacter de un coatí infectado con el que se les alimentó inadvertidamente (L. Saboia com. pers.). Esta pérdida de múltiples animales en un corto periodo de tiempo enfatiza tanto la sensibilidad de la especie a varias enfermedades como la susceptibilidad de las especies vivas del grupo a los patógenos. Con la creciente proximidad a las zonas humanas y la intrusión en zonas protegidas con perros de caza, el riesgo de exposición a enfermedades es potencialmente alto. La exposición a la enfermedad también puede deberse a los perros domésticos asilvestrados o semiasilvestrados, no vacunados, que cazan regularmente presas independientemente de los humanos (K. DeMatteo obs. pers.). DeMatteo (2008) señaló que en Brasil se observaron más interacciones problemáticas entre los perros de monte y los perros domésticos que con el ganado, lo que no hace sino enfatizar la amenaza de la enfermedad para la especie.
Información sobre el uso y el comercio
Los perros de monte han sido reportados por disparos en cinco países (Argentina, Brasil, Panamá, Perú, Venezuela) y atrapados en dos (Panamá y Perú) (DeMatteo 2008, F. Michalski datos no publicados). En Perú, estos eventos se asociaron con los nativos que informaron que comerán perros de monte si son atrapados o si tienen la rara oportunidad de recogerlos con la caza tradicional; sin embargo, los disparos reportados en Venezuela se asociaron con la recolección ocasional de trofeos debido a su rareza (DeMatteo 2008).
Información sobre acciones de conservación
Esta especie se encuentra en varias áreas protegidas a lo largo de su área de distribución, pero tiene una distribución irregular y ocurre en densidades muy bajas. La caza está prohibida en Colombia (Ley número 848:1973), Ecuador (Ley número 74:1981), Guayana Francesa (Ley número JO19860625:1986), Panamá (Ley número 2-80:1980), Paraguay (Ley número 18796:1975) y Perú (Ley número 5056:1970). La caza y el comercio están regulados en Argentina (Ley número 22.421:1981), Bolivia (Ley número 12301:1975), Brasil (Ley número 5197:191967) y Venezuela (Ley número 276:1970). No hay información sobre Guyana y Surinam. Lamentablemente, en muchas partes de su área de distribución, los recursos son inadecuados para gestionar las áreas protegidas designadas y hacer cumplir las leyes de vida silvestre existentes.
Incluido en CITES en el Apéndice I.
Los perros de monte se dan en cautividad y forman parte de un exitoso programa de cría internacional (Buck 2009), que incluye Asia (Japón), Europa, Brasil y América del Norte. No se conocen intentos de reintroducción.
Las estimaciones de población y los datos demográficos de los perros de monte son todavía poco conocidos en toda su área de distribución. Esto se extiende a la comprensión de la dinámica social de los grupos individuales, especialmente en términos de dispersión de las crías y el área de uso en relación con otros grupos (áreas de distribución superpuestas o separadas). Las asociaciones de hábitat no se comprenden claramente: antes se creía que la especie dependía de los bosques, pero ahora se observa cada vez más en hábitats abiertos y fragmentados; sin embargo, no hay datos sobre la viabilidad de la población en esas zonas. Aunque los datos preliminares sobre la dieta apoyan una dieta principalmente carnívora, es necesario evaluar los cambios estacionales y la variación geográfica de la dieta. Es necesario determinar cómo se puede gestionar o minimizar el impacto de las enfermedades, especialmente la dinámica de transmisión de los perros domésticos, en las poblaciones silvestres. Hay que seguir evaluando las relaciones interespecíficas con carnívoros simpátricos.
La localización de pruebas de la presencia de las especies mediante técnicas de estudio estándar, incluyendo cámaras trampa y estudios de transectos, ha resultado difícil (Beisiegel 2009, DeMatteo et al. 2009, Michalski 2010). El uso de señuelos de olor artificial, que pueden aumentar la atracción hacia un lugar específico, no ha tenido éxito con los perros de monte salvajes (Zuercher et al. 1999). Sin embargo, hay varios ajustes metodológicos que pueden aumentar la eficacia de estas técnicas con la especie, incluyendo el ajuste de la altura de la colocación de la cámara, el aumento del esfuerzo de trampeo y el uso simultáneo de reproducciones de vocalizaciones de llamadas largas y orina conespecífica (DeMatteo et al. 2004). Los ensayos de campo limitados en el cerrado parcialmente fragmentado (2004-2005) con playbacks, orina y trampas de sujeción de patas no tuvieron éxito (K. DeMatteo datos no publicados). Sin embargo, se necesitan ensayos adicionales para determinar cómo la densidad de las especies, la variabilidad del hábitat (bosque versus cerrado) y la densidad de las presas altera la efectividad de la técnica.
La investigación preliminar y en curso que utiliza una combinación de tres técnicas no invasivas (perros detectores de olor, detección de ADN fecal, tecnología GIS) ha demostrado ser exitosa para la especie (DeMatteo et al. 2009, datos inéditos) y debería ampliarse a otras regiones y hábitats. Este conjunto de técnicas elimina la necesidad de atraer a la especie a un lugar específico y permite conocer muchas variables ecológicas, incluyendo el uso del hábitat (intacto y fragmentado), el estado de la población, el área mínima de uso y el solapamiento/separación del nicho con otros carnívoros.
Leave a Reply