Frontiers in Plant Science
De forskellige typer af heterochroni
I 1870’erne identificerede Ernst Haeckel tidsmæssige og rumlige ændringer i udviklingen hos en efterkommer i forhold til forfaderen som de to vigtigste mekanismer for evolutionen (Haeckel, 1875). Haeckel kaldte de rumlige ændringer for heterotopi og de tidsmæssige ændringer for heterochroni.
Det er imidlertid sådan, at betydningen af heterochroni har ændret sig, siden Haeckel først opfandt begrebet. Haeckel brugte udtrykket heterochroni til at henvise til afvigelser fra hans velkendte “Biogenetiske Lov”, som siger, at rækkefølgen af udviklingshændelser (ontogeni) i vid udstrækning gentager rækkefølgen af begivenheder i artens udviklingshistorie (fylogeni) (Haeckel, 1875). Således henviste heterochroni oprindeligt til en ændring i tidspunktet for fremkomsten af et træk i en organismes udviklingssekvens i forhold til den sekvens, der opstod i organismens fylogeni. I midten af det 20. århundrede afkoblede De Beer (1951) heterochroni fra rekapitulation. Han brugte heterochroni til at betegne forskelle i timingen af udviklingshændelser, når man sammenligner to beslægtede arter, for at forklare, hvordan heterochroni kan skabe diversitet blandt organismer. Gould (1977) genforenede begrebet heterochroni med rekapitulation og definerede heterochroni som “ændringer i det relative tidspunkt for fremkomsten og udviklingshastigheden for karakterer, der allerede var til stede hos forfædrene”, idet han lagde vægt på ændringer i relativ størrelse og form snarere end i timingen af udviklingshændelser for at påvise heterochroni. Ved begyndelsen af det 21. århundrede foreslog Smith, at det ville være mere nyttigt at definere to typer heterochroni: “vækstheterochroni”, som lægger vægt på ændringer i den endelige størrelse og form, og “sekvensheterochroni”, som er tættere på Haeckels og de Beers oprindelige brug og giver mulighed for at forklare fænotypisk variation ved ændringer i timingen af udviklingshændelser (Smith, 2002, 2003; Keyte og Smith, 2014).
Sequence heterochrony (herefter benævnt ‘heterochrony’) kan inddeles i to kategorier: paedomorphosis og peramorphosis. Sammenlignet med forfædrenes udvikling resulterer paedomorfose i et juvenilt eller simpelt resultat, mens peramorfose resulterer i en voksen eller mere kompleks fænotype. Hver af disse to kategorier af heterokroni kan skyldes variation i timingen af starten, forskydningen eller hastigheden af en udviklingsproces, som foreslået af Alberch et al. (1979). Denne variation kan resultere i 6 forskellige typer af heterochroni (figur 1). Paedomorphosis kan skyldes en tidlig afslutning af en udviklingsproces (progenese), en forsinket start af processen (postdisposition) eller en langsommere udviklingshastighed (neoteny). Peramorfose er resultatet af en forlænget udviklingsperiode som følge af en senere afslutning (hypermorphose) eller en tidligere start (pre-displacement), eller af en højere udviklingshastighed (acceleration) (figur 1A). Hypotetiske eksempler på peramorfose og paedomorfose i planteembryogenese og vegetativ udvikling er vist i figur 1B,C.
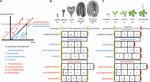
FIGUR 1. Typer af heterochroni. (A) Skematisk fremstilling af de 6 typer af heterochroni. Den sorte linje repræsenterer den tid, der er nødvendig for at nå et bestemt udviklingsstadium i den forfædres ontogeni. Blå linjer viser de 3 typer af pædomorfose: progenese (tidlig forskydning), postforskydning (forsinket begyndelse) og neoteni (langsommere udviklingshastighed). Røde linjer viser de 3 typer af peramorfose: hypermorphose (forsinket forskydning), pre-displacement (tidlig begyndelse) og acceleration (højere udviklingshastighed). Tegningen i (A) er baseret på Alberch et al. (1979) og Geuten og Coenen (2013). (B) Heterochrony-scenarier for embryogenese. Arabidopsis-embryogenese er her taget som den hypotetiske forfædres udvikling, opdelt i fire faser til illustrative formål. Et femte stadium, hvor embryoet har produceret de to første blade plus de to cotyledoner (betegnet embryonisk kimplante), foreslås som det sidste stadium i peramorf embryogenese, mens paedomorf embryogenese forventes at afsluttes på hjerte-stadiet. (C) Heterochroni-scenarier for vegetativ udvikling. I dette tilfælde repræsenterer Arabidopsis’ vegetative udvikling, hvor planten fortsætter med at producere cauline/reproduktive blade på stænglen før blomstring (tegnet øverst), et hypotetisk tilfælde af peramorf vegetativ udvikling, sammenlignet med en hypotetisk forfader, der blomstrer i voksenstadiet uden at producere caulineblade. Paedomorf vegetativ udvikling forudsiges at resultere i planter, der blomstrer på ungdomsstadiet.
Mens disse klassifikationer er nyttige til at illustrere ændringer i udviklingstidspunktet, er det ofte vanskeligt at skelne mellem forskellige former for heterochroni, når man ser på eksempler fra det virkelige liv. I mange tilfælde kan evolutionen resultere i forskellige typer heterokroni, der hver især forekommer på et diskret stadium af en udviklingsproces (gennemgået i planter af Li og Johnston, 2000).
Heterokroni i planternes udvikling og diversificering
Landplanter (embryofytter) har gennemgået mange morfologiske innovationer siden deres opståen i midten af den ordoviske periode for ca. 470 millioner år siden. Tidlige divergerende slægter opstod i løbet af en periode på mere end 100 millioner år i løbet af silur og tidlig devon (Kenrick og Crane, 1997; Pires og Dolan, 2012; Harrison og Morris, 2018). Embryofytter udviklede sig fra en forfader af ferskvandsalger, der var beslægtet med de nuværende charofytgrupper Charales, Coleochaetales og Zygnematales. Overgangen til vækst på land, som observeret i bryofytter (hornvort, mosser og levermosser), involverede oprindelsen af sporer, vekslen af gametofyt- og sporofytgenerationer, ensidige former og tredimensionel vækst. Innovationer hos terrestriske karplanter (lycofytter, monilofytter og spermatofytter) omfattede bifurkation, ubestemthed, sporofytisk dominans, axillær forgrening og dannelse af meristemer, blade og rødder (Harrison, 2017).
Flere af de ovennævnte evolutionære trin er blevet knyttet til heterochroni. For oprindelsen af sporogenese tyder data fra bryofyter på, at sporer blev produceret direkte fra zygoter i en proces, der involverer tidlig cytokinesis, acceleration af meiose og forsinket vægdeponering fra zygoten til meiosporerne (Brown og Lemmon, 2011). Den forgrenede sporofyt (polisporangiofyt) er blevet antaget at have udviklet sig ved udvidet vegetativ vækst af den apikale celle. Denne længere periode med vegetativ vækst blev foreslået at resultere i en forlænget embryonal akse, skudforgrening og en forsinkelse i overgangen til reproduktiv vækst, der producerer sporangiet (Rothwell et al., 2014; Tomescu et al., 2014). Yderligere undersøgelser af plantefossiler skulle give flere beviser for gamle plantemorfologier, hvilket ville gøre det muligt at sammenligne nutidige og uddøde former (Rothwell et al., 2014).
I uddøde planter er der blevet identificeret heterokroniske ændringer i gametofytudvikling, embryogenese, vegetativ udvikling, skudmodning og blomstermorfogenese. Den kvindelige gametofyt af Gnetum er strukturelt afvigende fra andre planter på grund af forskelle i timingen af befrugtning og somatisk udvikling. Tidsmæssige ændringer i cellecyklusprogressionen har bidraget til diversificerede tidsmønstre for spermatogenese og gametfusion under befrugtning (Friedman, 1999; Tian et al., 2005). I et tilfælde af progenese sker befrugtning på et frit nukleært stadium af somatisk udvikling, et juvenilt stadium sammenlignet med den forfædres somatiske ontogeni, hvilket udelukker differentiering af ægceller (Friedman og Carmichael, 1998). Den apomiktiske udvikling af Boechera ægceller er blevet forbundet med heterokroniske genekspressionsmønstre sammenlignet med ikke-apomiktiske (seksuelle) ægceller (Sharbel et al., 2010). Udviklingen af Rafflesiaceae, en holoparasitisk plantefamilie, som inficerer vinplanter, viser to heterokroniske skift: et stop på proembryonstadiet, som kan betragtes som et eksempel på neoteni, og en acceleration af overgangen fra den udifferentierede endofyt til blomstring, idet den vegetative skudmodning springes over (Nikolov et al, 2014).
En QTL-analyse (Quantitative Trait Loci), der sammenligner Eucalyptus globulus-populationer med tidlig vegetativ faseændring og populationer, hvor vegetativ faseændring er forsinket flere år, identificerede ekspressionen af mikroRNA’et EglMIR156.5 som ansvarlig for heterokronisk variation i vegetativ faseændring i E. globulus (Hudson et al., 2014). En anden QTL-analyse konkluderede, at heterokroni ligger til grund for den naturlige variation i Cardamine hirsuta bladform (Cartolano et al., 2015). QTL-kortlægning fastslog, at effekten skyldes cis-regulerende variation i blomsterrepressoren ChFLC, således at populationer med lavt eksprimerende ChFLC-alleler viser både tidlig blomstring og fremskyndet erhvervelse af voksne bladegenskaber, især øget bladpletantal. Morfometriske og QTL-analyser har fastslået, at heterokroniske mutationer bidrager til den naturlige variation i Antirrhinum og til druehøjblomstring af druer (Costa et al., 2012; Chitwood et al., 2016). En Principal Component Analysis (PCA) af de ontogenetiske baner af bladform blandt de tre slægter af marsileaceous bregner (Marsilea, Regnellidium og Pilularia) foreslog, at de viser en paedomorfisk fænotype, sammenlignet med den mere komplekse forfædres udvikling, forårsaget af accelereret væksthastighed og tidlig afslutning på en forenklet bladform (Pryer og Hearn, 2009).
Evolutionær diversitet i blomsterstandens arkitektur hos Solanaceae er moduleret af heterokroniske skift i erhvervelse af blomstringsskæbne (Lippman et al, 2008; Park et al., 2012). En sammenligning af transkriptomer af meristemmodning fra fem domesticerede og vilde Solanaceae-arter afslørede et peak af ekspressionsdivergens, der ligner “omvendt timeglas”-modellen for dyreembryogenese, som siger, at en midt i udviklingsperioden af divergens driver morfologisk variation (Lemmon et al., 2016). I græsser resulterer en forsinkelse i overgangen fra skudmeristem (SM) til blomstermeristem (FM) i mere komplekse panikler (Kyozuka et al., 2014). Popler (Populus sp.) og pil (Salix sp.) bærer kompakte unisexuelle blomsterstande kendt som “catkins”, som er blevet foreslået at være udviklet ud fra en forenkling af panikformen ved en tidlig SM- til FM-overgang (Cronk et al., 2015). Heterokroniske ændringer har også bidraget til naturlig variation i blomstringstidspunkt og skudarkitektur blandt Mimulus guttatus-populationer (Baker og Diggle, 2011).
Morfologisk diversitet af perianth i Dipsacoideae er forårsaget af heterokroniske ændringer i organinitiering, specifikt i antallet af sepaler (Naghiloo og Claßen-Bockhoff, 2017). Den store formdiversitet af sepaler blandt Iris-arter skyldes mere heterochroni end heterotopiske ændringer (Guo, 2015). En undersøgelse i Brassicaceae viste, at evolutionen af corolla monosymmetri fra den polysymmetriske ancestrale blomst involverede et heterokronisk skift i ekspressionen af CYC2 gener (en klade af TCP transkriptionsfaktorer) fra tidlig adaxial ekspression i ancestrale blomstermeristemer, til en senere adaxial ekspression i kronbladets udvikling (Busch et al., 2012). Heterochronisk, men ikke heterotopisk, CYC2-ekspression er også blevet forbundet med et tab af papillate kegleformede celler i kronblade og et skift til fuglebestøvningssystem i Lotus (Ojeda et al., 2017). En paedomorphisk morfologi, hvor blomsterne holder modne pollen i uåbnede knoppelignende strukturer, førte til specialiseret bestøvning i en klade af Madagaskar-vinplanter (Euphorbiaceae) (Armbruster et al., 2013). Udviklingen af cleistogame capitulum fra en chasmogamous ancestral tilstand er et klassisk eksempel på paedomorphosis, da den cleistogame form viser juvenile træk (Lord og Hill, 1987). Cleistogami hos Asteraceae udviklede sig specifikt ved præ-forskydning og progenese af blomsterudviklingen samt neoteni af alle andre hvirvler end gynoeciumet (Porras og Muñoz, 2000). Mangfoldigheden af blomstermorfologier inden for Jaltomata, en Solanaceae-slægt, skyldes hypermorphose og acceleration af nogle corolla-egenskaber (Kostyun et al., 2017). Diversiteten af Azoriske sommerfugleorkidéer skyldes også florale heterokroniske skift (Bateman et al., 2014). For nylig understregede Ronse de Craene (2018) betydningen af heterochroni i tre udviklingsprocesser: phyllotaxis, udviklingen af fælles støvfangs- og blomsterprimordier og obdiplostemony, idet han forbandt ændringer i væksthastigheden med forsinket organinitiering. Heterochroniske væksthastigheder af perianth og stil og tidlig hypanthiumforlængelse er ansvarlige for den store artsdiversitet inden for den morfologisk homogene Eugenia-slægt (Vasconcelos et al., 2018). Endelig er heterokronisk udtryk af fw2.2-allelen, som påvirker celledeling i tidlig frugtudvikling, ansvarlig for naturlig variation i tomatfrugtstørrelse (Cong et al., 2002).
Studying Heterochronic Mutants to Elucidate Genetic Control of Timing
Studiet af mutanter, der er påvirket i udviklingstiming, har kastet lys over genetiske veje, der kontrollerer morfogenese og udviklingsmæssige overgange. Heterochroni kan være forårsaget af tidligere eller senere aktivering eller undertrykkelse af disse veje.
Lavey cotyledon (lec), dicer-like1 (dcl1) og ekstra cotyledon (xtc) mutanter af Arabidopsis thaliana repræsenterer heterochroniske fænotyper, der har hjulpet med at definere frømodningsprogrammer. lec mutanter producerer cotyledoner med træk af bladidentitet (Meinke, 1992), et klart eksempel på homeose: udskiftning af en struktur med en anden. Det er imidlertid ofte vanskeligt at skelne mellem homeose og heterochroni, da homeose kan være resultatet af både heterochroni og heterotopi (Li og Johnston, 2000; Geuten og Coenen, 2013). Under sen embryogenese fremmer LEC2 frømodning og undertrykker postembryonal identitet (Stone et al., 2008). Ligesom lec-mutanter viser dcl1-mutanter peramorfe fænotyper under embryogenese, da kloroplastudvikling og frøopbevaringsprotein-genekspression forekommer tidligere end i wild type-embryoner. DCL1 er påkrævet for biogenese af mikroRNA’er, som undertrykker frømodning gennem masterregulatorerne LEC2 og FUSCA3 (Nodine og Bartel, 2010; Willmann et al., 2011). xtc1-, xtc2- og altered meristem programming1 (amp1)-mutanterne viser en homeotisk fænotype, hvor de første et eller to blade omdannes til cotyledoner (Conway og Poethig, 1997). I disse tre mutanter er den kugleformede overgang til hjerteovergangen forsinket, hvilket medfører et forstørret skudmeristem, som fører til ekstra organdannelse under embryogenesen. Denne fænotype kan fortolkes som hypermorfose, en type peramorfose, da embryonets udvikling udvides til en mere udviklet form (ligesom det peramorfe scenario i figur 1B). Hvis den vegetative udvikling imidlertid vælges som referencepunkt, ville lec-mutanter repræsentere et tilfælde af præ-displacement (peramorfose), og xtc1, xtc2 og amp1 ville repræsentere post-displacement (paedomorphosis) i erhvervelse af bladidentitet. AMP1 er påkrævet for miRNA-medieret translationel repression på ER-membranen (Li et al., 2013).
Genetisk regulering af den juvenile til voksen overgang, også kaldet vegetativ faseændring, er blevet undersøgt ved hjælp af både paedomorfe og peramorfe mutanter. Screeninger for mutanter, der viser en tidlig voksen (peramorfe) fænotype, producerede alleler af gener relateret til små RNA-biogenese som zippy/ago7, sgs3 og rdr6 (Hunter et al., 2003; Peragine et al., 2004), mens mutanter med en sen voksen (paedomorfe) fænotype skyldes øgede miRNA-niveauer (Gillmor et al., 2014; Xu M. et al., 2016, 2018; Guo et al., 2017). Det genetiske grundlag for disse fænotyper forklares ved ændret ekspression af de nært beslægtede mikroRNA’er miR156 og miR157, de vigtigste regulatorer af vegetativ faseændring, der virker ved at undertrykke SPL-transskriptionsfaktorer på både transkriptionelt og translationelt niveau på en tærskelafhængig måde (Wu og Poethig, 2006; Chuck et al., 2007; Poethig, 2013; He et al., 2018). Både MIR156- og SPL-genfamilier er direkte reguleret af epigenetiske mærker: MIR156 aktiveres af H3K4me3 og H3K4ac, som fremmes af SWR1-C-komplekset og kromatin remodeler BRAHMA, og undertrykkes af H3K27me3, som fremmes af Polycomb-proteinerne SWINGER og CURLY LEAF og af kromatin remodeler PICKLE (Xu M. et al., 2016; Xu Y. et al., 2016; Xu M. et al., 2018). SPL-gener aktiveres af histonacetylering medieret af det SAGA-lignende kompleks og undertrykkes af H2AUb medieret af Polycomb-proteinerne RING1A og RING1B (Kim et al., 2015; Li et al., 2017). Undersøgelsen af planter med tab- eller gain-of-funktion af SPL-gener har også defineret en endogen blomstringsvej, hvor nogle SPL-gener fremmer ekspressionen af miR172, som igen fremmer blomstring ved at undertrykke APETALA2-familiens blomstringsrepressorer (Wang et al., 2009; Wu et al., 2009; Huijser og Schmid, 2011).
Suden større udviklingsovergange kan organmorfogenese også påvirkes af heterokronisk aktivering af genetiske regulatorer. For eksempel resulterer tidlig spiring af Brassica rapa-embryoner i organer med mosaikker af cotyledon- og bladidentitet (Fernandez, 1997), og differentiel tidsmæssig ekspression af klasse II af TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATION CELL FACTOR (TCP)-generne resulterer i blade med forskellig størrelse og form (Efroni et al, 2008).
Mekanismer, der driver ændringer i udviklingstiming i planter (transkriptionel, metabolisk og cellulær heterochroni)
Transkriptionel heterochroni henviser til en ændring i timingen af aktivering eller undertrykkelse af genekspression og er ofte forårsaget af ændringer i cis-regulerende genområder (Pham et al., 2017). Transkriptionel heterochroni i de ovenfor nævnte genetiske veje er en almindelig måde at producere heterochroniske fænotyper på. Metabolisk kontrol af veje, der regulerer udviklingsovergange (her benævnt “metabolisk heterochroni”), og tidsmæssig kontrol af celleproliferation, celleudvidelse og celledifferentiering (her benævnt “cellulær heterochroni”) er andre mekanismer, der driver heterochroni i planter (figur 2). Transkriptionelle, metaboliske og cellulære processer er indbyrdes forbundne, så den molekylære oprindelse af heterochroni kan skyldes en kombination af flere mekanismer.

FIGUR 2. Mekanismer, der driver heterochroni i planter. (A) Cellulær heterochroni. Rækkefølgen af rosetblade, der produceres under den vegetative udvikling, er vist. Fænotypen af Arabidopsis Columbia-wildtypeplanter er taget som en hypotetisk forfødt tilstand; et paedomorft scenarie (neoteni) og et peramorft scenarie (acceleration) er også vist. Pilene repræsenterer retningen af bølgen af celledelingsstop i en basipetal gradient, hvor cellerne under den stiplede linje stadig deler sig aktivt, mens cellerne ovenover udvider sig og/eller differentierer sig. (B) Transkriptionel og metabolisk heterochroni. En grafisk repræsentation af den forudsagte hyppighed af mikroRNA miR156/7 og dets mål, SPL-generne, samt sukkerhyppigheden langs den vegetative udviklingstid, svarende til fænotyperne i (A). De stiplede linjer repræsenterer den tærskel for miR156/7- og SPL-overflod, der fører til ungdomsstadiet (over tærsklen) eller voksenstadiet (under tærsklen). I dette tilfælde resulterer en forsinkelse i repressionen af miR156 og aktivering af SPL-ekspression og sukkerproduktion i paedomorfose, mens en tidlig nedbrydning af miR156 og en tidlig aktivering af SPL-ekspression og sukkerproduktion resulterer i peramorfose. Disse tegninger er forenklede gengivelser og afspejler ikke de nøjagtige forekomster i naturen. Det faktiske mønster af miR156/SPL-overflod er tættere på det afbildede for “peramorfose”-tilstand.
Metabolisk heterochroni kan påvirkes af hormoner, sukkerstoffer og redox-signaler (Jia et al., 2017). Differentiel biosyntese, transport og perception af hormoner såsom auxin, jasmonsyre (JA), gibberellin og abscisinsyre påvirker heterochroni ved at kontrollere regulatorer af udviklingsprocesser. For eksempel forsinker mutanter i AUXIN RESPONSE FACTORS (ARF) ARF3 og ARF4 den voksne overgang (Fahlgren et al., 2006; Hunter et al., 2006), og auxin homeostase kontrollerer overgangen fra vedligeholdelse af blomsterstamceller til gynoeciumdannelse (Yamaguchi et al., 2017). Eksogen JA kan forsinke den voksne overgang ved at udskyde faldet i miR156-ekspressionen (Beydler et al., 2016), og gibberellin fremskynder blomstringen ved at frigøre SPL-gener fra repression af DELLA-proteiner (Yu et al., 2012). Ernæringsstatus har været forbundet med kontrollen af vegetativ faseændring siden begyndelsen af det 20. århundrede (Goebel, 1908). Sukker produceret af fotosyntese er nødvendigt for erhvervelse af voksne egenskaber og er delvist ansvarlig for faldet i miR156-ekspressionen i sen vegetativ udvikling (Yang et al., 2013; Yu et al., 2013; Buendía-Monreal og Gillmor, 2017). HEXOKINASE1 (HXK1) og Trehalose-6-fosfat (T6P) er vigtige for den sukkermedierede repression af miR156 og fremmer derved vegetativ faseændring og blomstring (Wahl et al., 2013; Yang et al., 2013).
På celleniveau består organogenese af en sekvens af tre faser: etablering af polaritet, celleproliferation og celleudvidelse (Walcher-Chevillet og Kramer, 2016). Timingen af initiering og afslutning af disse stadier er afgørende for organernes størrelse og form, og heterochroni i denne sekvens resulterer i diversificering af organstørrelse og -form (Figur 2). Overgangen fra celleproliferation til celleudvidelse og differentiering kræver koordinering mellem cellecyklus og cellevækst (Sablowski og Carnier Dornelas, 2014). I blade af modelplanter bevæger denne overgang sig som en basipetal bølge af cellecyklusstop, der begynder ved den distale del af primordiet og bevæger sig mod basen. Cellerne bag mitotisk arrestfronten bliver stærkt vacuoliserede og begynder at ekspandere (Donnelly et al., 1999; Czesnick og Lenhard, 2015). Andre plantearter kan imidlertid vise diffus vækst og acropetale eller bidirektionale cellecyklusstopgradienter (Das Gupta og Nath, 2015). Erhvervelse af fotosyntetisk kapacitet er nødvendig for skiftet fra celledeling til celleudvidelse (Andriankaja et al., 2012). Dette skift korrelerer med sukkerets rolle med hensyn til at fremme Target of Rapamycin (TOR)-vejen og undertrykke Sucrose-non-fermenting1-relateret kinase 1 (SnRK1): TOR og T6P inducerer celleudvidelse ved at fremme makromolekylær syntese, mens SnRK1 fremmer katabolisme (Tsai og Gazzarrini, 2014; Sablowski, 2016). To mikroRNA’er spiller modsatte roller i dette cellulære skift: miR319 undertrykker ekspressionen af klasse II TCP-faktorer, som er hæmmere af celleproliferation, mens miR396 begrænser ekspressionen af GROWTH REGULATING FACTORS (GRF’er), som forsinker differentiering (Das Gupta og Nath, 2015; Maugarny-Calés og Laufs, 2018). Overgangen fra et ubestemt shoot apical meristem til et bestemt blomstermeristem involverer også tidsmæssig regulering af den cellulære identitet. Timingen af AGAMOUS aktivering af KNUCKLES, som igen undertrykker WUSCHEL, definerer det tidsmæssige vindue for ubestemthed og dermed størrelsen og antallet af organer (Sun et al., 2014).
Slutning
En forståelse af tidsmæssig regulering af planteudvikling er nødvendig for bedre at kunne værdsætte den mangfoldighed af planteformer, som vi ser i naturen, for at forklare planternes morfologiske evolution og for at kunne manipulere plantearkitekturen til gavn for landbruget. Som skitseret ovenfor styrer indbyrdes forbundne transkriptionelle, metaboliske og cellulære mekanismer heterochroni i eksisterende arter. Yderligere forskning i disse veje i angiospermer og basale planteafstamninger bør afsløre mere om de ændringer i udviklingstidspunktet, der har drevet udviklingen af udvikling i planter.
Author Contributions
MB-M udtænkte og skrev manuskriptet. CSG redigerede og reviderede manuskriptet.
Interessekonflikt erklæring
Forfatterne erklærer, at forskningen blev udført uden kommercielle eller økonomiske relationer, der kunne opfattes som en potentiel interessekonflikt.
Alberch, P., Gould, S., Oster, G., og Wake, D. (1979). Størrelse og form i ontogeni og fylogeni. Paleobiology 5, 296-317. doi: 10.1017/S0094837300006588
CrossRef Full Text | Google Scholar
Andriankaja, M., Dhondt, S., DeBodt, S., Vanhaeren, H., Coppens, F., DeMilde, L., et al. (2012). Udgang fra proliferation under bladudvikling i Arabidopsis thaliana: en ikke-så-graduel proces. Dev. Cell 22, 64-78. doi: 10.1016/j.devcel.2011.11.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Armbruster, W. S., Lee, J., Edwards, M. E., og Baldwin, B. G. (2013). Floral paedomorfi fører til sekundær specialisering i bestøvning af Madagaskar Dalechampia (Euphorbiaceae). Evolution 67, 1196-1203. doi: 10.1111/evo.12002
PubMed Abstract | CrossRef Full Text | Google Scholar
Baker, R. L., og Diggle, P. K. (2011). Knude-specifik forgrening og heterokroniske ændringer ligger til grund for forskelle på populationsniveau i Mimulus guttatus (Phrymaceae) skudarkitektur. Am. J. Bot. 98, 1924-1934. doi: 10.3732/ajb.1100098
PubMed Abstract | CrossRef Full Text | Google Scholar
Bateman, R. M., Rudall, P. J., Bidartondo, M. I., Cozzolino, S., Tranchida-Lombardo, V., Carine, M. A., et al. (2014). Speciation via blomstret heterochroni og formodet mykorrhizal værtsskifte af endemiske sommerfugleorkidéer på Azorernes øgruppe. Am. J. Bot. 101, 979-1001. doi: 10.3732/ajb.1300430
PubMed Abstract | CrossRef Full Text | Google Scholar
Beydler, B. D., Osadchuk, K., Cheng, C.-L., Manak, J. R., og Irish, E. E. (2016). I majsens juvenile fase ses opregulering af stressresponsgener og forlænges af eksogen JA. Plant Physiol. 171, 2648-2658. doi: 10.1104/pp.15.01707
PubMed Abstract | CrossRef Full Text | Google Scholar
Brown, R. C., og Lemmon, B. E. (2011). Sporer før sporofytter: hypoteser om oprindelsen af sporo- genese ved overgangen mellem alger og planter. New Phytol. 190, 875-881. doi: 10.1111/j.1469-8137.2011.03709.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Buendía-Monreal, M., og Gillmor, C. S. (2017). Konvergent repression af miR156 af sukker og CDK8-modulet af Arabidopsis Mediator. Dev. Biol. 423, 19-23. doi: 10.1016/j.ydbio.2017.01.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Busch, A., Horn, S., Mühlhausen, A., Mummenhoff, K., og Zachgo, S. (2012). Corolla monosymmetri: evolution af en morfologisk nyhed i Brassicaceae-familien. Mol. Biol. Evol. 29, 1241-1254. doi: 10.1093/molbev/msr297
PubMed Abstract | CrossRef Full Text | Google Scholar
Cartolano, M., Pieper, B., Lempe, J., Tattersall, A., Huijser, P., Tresch, A., et al. (2015). Heterochrony underpins natural variation i Cardamine hirsuta bladform. Proc. Natl. Acad. Sci. U.S.A. 112, 10539-10544. doi: 10.1073/pnas.1419791112
PubMed Abstract | CrossRef Full Text | Google Scholar
Chitwood, D. H., Klein, L. L., O’Hanlon, R., Chacko, S., Greg, M., Kitchen, C., et al. (2016). Latente udviklingsmæssige og evolutionære former indlejret i vindruebladet. New Phytol. 210, 343-355. doi: 10.1111/nph.13754
PubMed Abstract | CrossRef Full Text | Google Scholar
Chuck, G., Cigan, A. M., A. M., Saeteurn, K., og Hake, S. (2007). Den heterokroniske majsmutant corngrass1 skyldes overekspression af et tandem microRNA. Nat. Genet. 39, 544-549. doi: 10.1038/ng2001
PubMed Abstract | CrossRef Full Text | Google Scholar
Cong, B., Liu, J., og Tanksley, S. D. (2002). Naturlige alleler på et kvantitativt egenskabslocus for tomatfrugternes størrelse adskiller sig ved heterokroniske regulerende mutationer. Proc. Natl. Acad. Sci. U.S.A. 99, 13606-13611. doi: 10.1073/pnas.172520999
PubMed Abstract | CrossRef Full Text | Google Scholar
Conway, L. J., og Poethig, R. S. (1997). Mutationer af Arabidopsis thaliana, der omdanner blade til cotyledoner. Proc. Natl. Acad. Sci. U.S.A. 94, 10209-10214. doi: 10.1073/pnas.94.19.10209
PubMed Abstract | CrossRef Full Text | Google Scholar
Costa, M. M. M. R., Yang, S., Critchley, J., Feng, X., Wilson, Y., Langlade, N., et al. (2012). Det genetiske grundlag for naturlig variation i heteroblasty i Antirrhinum. New Phytol. 196, 1251-1259. doi: 10.1111/j.1469-8137.2012.04347.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Cronk, Q. C. B., Needham, I., og Rudall, P. J. (2015). Evolution af kattekillinger: blomsterstandsmorfologi hos udvalgte Salicaceae i en evolutionær og udviklingsmæssig sammenhæng. Front. Plant Sci. 6:1030. doi: 10.3389/fpls.2015.01030
PubMed Abstract | CrossRef Full Text | Google Scholar
Czesnick, H., og Lenhard, M. (2015). Størrelseskontrol i planter – lektioner fra blade og blomster. Cold Spring Harb. Perspect. Biol. 7:a019190. doi: 10.1101/cshperspect.a019190
PubMed Abstract | CrossRef Full Text | Google Scholar
Das Gupta, M., and Nath, U. (2015). Divergens i mønstre af bladvækstpolaritet er forbundet med udtryksdivergens af miR396. Plant Cell 27, 2785-2799. doi: 10.1105/tpc.15.00196
PubMed Abstract | CrossRef Full Text | Google Scholar
De Beer, G. R. (1951). Embryoner og forfædre. Oxford: Clarendon Press.
Google Scholar
Donnelly, P. M., Bonetta, D., Tsukaya, H., Dengler, R. E., og Dengler, N. G. (1999). Cellecyklus og celleforøgelse i udviklende blade af Arabidopsis. Dev. Biol. 215, 407-419. doi: 10.1006/dbio.1999.9443
PubMed Abstract | CrossRef Full Text | Google Scholar
Efroni, I., Blum, E., Goldshmidt, A., og Eshed, Y. (2008). Et langvarigt og dynamisk modningsskema ligger til grund for Arabidopsis’ bladudvikling. Plant Cell 20, 2293-2306. doi: 10.1105/tpc.107.057521
PubMed Abstract | CrossRef Full Text | Google Scholar
Fahlgren, N., Montgomery, T. A., Howell, M. D., Allen, E., Dvorak, S. K., Alexander, A. L., et al. (2006). Regulering af AUXIN RESPONSE FACTOR3 af TAS3 ta-siRNA påvirker udviklingstidspunktet og mønsterdannelsen i Arabidopsis. Curr. Biol. 16, 939-944. doi: 10.1016/j.cub.2006.03.065
PubMed Abstract | CrossRef Full Text | Google Scholar
Fernandez, D. E. (1997). Udviklingsgrundlaget for homeose i tidligt spirende Brassica napus-embryoner: faseændring ved skudspidsen. Development 124, 1149-1157.
PubMed Abstract | Google Scholar
Friedman, W. E. (1999). Ekspression af cellecyklus i sædceller fra Arabidopsis: implikationer for forståelsen af mønstre for gametogenese og befrugtning i planter og andre eukaryoter. Development 126, 1065-1075.
PubMed Abstract | Google Scholar
Friedman, W. E., og Carmichael, J. S. (1998). Heterochroni og udviklingsmæssig innovation: evolution af kvindelig gametofytontogeni i gnetum, en stærkt apomorphisk frøplante. Evolution 52, 1016-1030. doi: 10.1111/j.1558-5646.1998.tb01830.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Geuten, K., and Coenen, H. (2013). Heterochroniske gener i planters evolution og udvikling. Front. Plant Sci. 4:381. doi: 10.3389/fpls.2013.00381
PubMed Abstract | CrossRef Full Text | Google Scholar
Gillmor, C. S., Silva-Ortega, C. O., Willmann, M. R., Buendía-Monreal, M., og Poethig, R. S. (2014). Arabidopsis Mediator CDK8-modulgenerne CCT (MED12) og GCT (MED13) er globale regulatorer af udviklingsfaseovergange. Development 141, 4580-4589. doi: 10.1242/dev.111229
PubMed Abstract | CrossRef Full Text | Google Scholar
Goebel, K. (1908). Einleitung in die Experimentelle Morphologie der Pflanzen. Leipzig: B.G. Teubner.
Google Scholar
Gould, S. J. (1977). Ontogeny and Phylogeny. Cambridge, MA: Harvard University Press.
Google Scholar
Guo, C., Xu, Y., Shi, M., Lai, Y., Wu, X., Wang, H., et al. (2017). Repression af miR156 af miR159 regulerer timingen af overgangen fra juvenil til voksen i Arabidopsis. Plant Cell 29, 1293-1304. doi: 10.1105/tpc.16.00975
PubMed Abstract | CrossRef Full Text | Google Scholar
Guo, J. (2015). Mekanismer for evolutionen af komplekse og mangfoldigt udarbejdede sepaler i Iris identificeret ved sammenlignende analyse af udviklingssekvenser. Am. J. Bot. 102, 819-832. doi: 10.3732/ajb.1400519
PubMed Abstract | CrossRef Full Text | Google Scholar
Haeckel, E. (1875). Die Gastrula und die Eifurchung der Thiere. Jena. Z. Naturwiss. 9, 402-508.
Google Scholar
Harrison, C. J. (2017). Udvikling og genetik i udviklingen af landplanters kropsplaner. Philos. Trans. R. Soc. B Biol. Sci. 372:20150490. doi: 10.1098/rstb.2015.0490
PubMed Abstract | CrossRef Full Text | Google Scholar
Harrison, C. J., og Morris, J. L. (2018). Oprindelsen og den tidlige evolution af karplanters skud og blade. Philos. Trans. R. Soc. B Biol. Sci. 373:20160496. doi: 10.1098/rstb.2016.0496
PubMed Abstract | CrossRef Full Text | Google Scholar
He, J., Xu, M., Willmann, M. R., Mccormick, K., Hu, T., Yang, L., et al. (2018). Tærskelafhængig repression af SPL-genekspression af miR156/miR157 kontrollerer vegetativ faseændring i Arabidopsis thaliana. PLoS Genet. 14:e1007337. doi: 10.1371/journal.pgen.1007337
PubMed Abstract | CrossRef Full Text | Google Scholar
Hudson, C. J., Freeman, J. S., Jones, R. C., Potts, B. M., Wong, M. M. L., Weller, J. L., et al. (2014). Genetisk kontrol af heterochroni i Eucalyptus globulus. G3 4, 1235-1245. doi: 10.1534/g3.114.011916
PubMed Abstract | CrossRef Full Text | Google Scholar
Huijser, P., og Schmid, M. (2011). Kontrol af udviklingsfaseovergange i planter. Development 138, 4117-4129. doi: 10.1242/dev.063511
PubMed Abstract | CrossRef Full Text | Google Scholar
Hunter, C., Sun, H., og Poethig, R. S. (2003). Arabidopsis heterochronic-genet ZIPPY er et medlem af ARGONAUTE-familien. Curr. Biol. 13, 1734-1739. doi: 10.1016/j.cub.2003.09.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Hunter, C., Willmann, M. R., Wu, G., Yoshikawa, M., de la Luz Gutiérrez-Nava, M., og Poethig, R. S. (2006). Trans-virkende siRNA-medieret repression af ETTIN og ARF4 regulerer heteroblasty i Arabidopsis. Development 133, 2973-2981. doi: 10.1242/dev.02491
PubMed Abstract | CrossRef Full Text | Google Scholar
Jia, X. L., Chen, Y. K., Xu, X. Z., Shen, F., Zheng, Q. B., Du, Z., et al. (2017). miR156 skifter på vegetativ faseændring under regulering af redox-signaler i æblefrøplanter. Sci. Rep. 7, 1-13. doi: 10.1038/s41598-017-14671-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Kenrick, P., og Crane, P. R. (1997). Oprindelsen og den tidlige udvikling af planter på land. Nature 389, 33-39. doi: 10.1038/37918
CrossRef Full Text | Google Scholar
Keyte, A. L., og Smith, K. K. (2014). Heterochroni og udviklingstidsmekanismer: skiftende ontogenier i evolutionen. Semin. Cell Dev. Biol. 34, 99-107. doi: 10.1016/j.semcdb.2014.06.015
PubMed Abstract | CrossRef Full Text | Google Scholar
Kim, J. Y., Oh, J. E., Noh, Y. S., og Noh, B. (2015). Epigenetisk kontrol af juvenil til voksen faseovergang af det Arabidopsis SAGA-lignende kompleks. Plant J. 83, 537-545. doi: 10.1111/tpj.12908
PubMed Abstract | CrossRef Full Text | Google Scholar
Kostyun, J. L., Preston, J. C., og Moyle, L. C. (2017). Heterokroniske udviklingsmæssige skift ligger til grund for blomsterdiversitet inden for Jaltomata ( Solanaceae ). Evodevo 8:17. doi: 10.1186/s13227-017-0080-z
PubMed Abstract | CrossRef Full Text | Google Scholar
Kyozuka, J., Tokunaga, H., og Yoshida, A. (2014). Kontrol af græsblomsterstandens form ved finjustering af meristemfaseskift. Curr. Opin. Plant Biol. 17, 110-115. doi: 10.1016/j.pbi.2013.11.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Lemmon, Z. H., Park, S. J., Jiang, K., Van Eck, J., Schatz, M. C., og Lippman, Z. B. (2016). Udviklingen af blomsterstandens diversitet i natskygge og heterochroni under meristemmodning. Genome Res. 26, 1676-1686. doi: 10.1101/gr.207837.116
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, J., Wang, Z., Hu, Y., Cao, Y., og Ma, L. (2017). Polycomb-gruppeproteinerne RING1A og RING1B regulerer den vegetative faseovergang i Arabidopsis. Front. Plant Sci. 8:867. doi: 10.3389/fpls.2017.00867
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, P., og Johnston, M. O. (2000). Heterochroni i planteevolutionsstudier gennem det tyvende århundrede. Bot. Rev. 66, 57-88. doi: 10.1007/BF02857782
CrossRef Full Text | Google Scholar
Li, S., Liu, L., Zhuang, X., Yu, Y., Liu, X., Cui, X., et al. (2013). MicroRNA’er hæmmer oversættelsen af mål-mRNA’er på det endoplasmatiske reticulum i Arabidopsis. Cell 153, 562-574. doi: 10.1016/j.cell.2013.04.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Lippman, Z. B., Cohen, O., Alvarez, J. P., Abu-Abied, M., Pekker, I., Paran, I., et al. (2008). Oprettelse af en sammensat blomsterstand i tomat og beslægtede natskyggeplanter. PLoS Biol. 6:e288. doi: 10.1371/journal.pbio.0060288
PubMed Abstract | CrossRef Full Text | Google Scholar
Lord, E. M., og Hill, J. P. (1987). “Evidence for heterochrony in the evolution of plant form,” in Development as an Evolutionary Process, eds R. A. Raff and E. C. Raff (New York, NY: Alan R. Liss), 47-70.
Google Scholar
Maugarny-Calés, A., and Laufs, P. (2018). At få blade i form: et molekylært, cellulært, miljømæssigt og evolutionært synspunkt. Development 145:dev16161646. doi: 10.1242/dev.161646
PubMed Abstract | CrossRef Full Text | Google Scholar
Meinke, D. W. (1992). En homoetisk mutant af Arabidopsis thaliana med bladformede cotyledoner. Science 258, 1647-1649. doi: 10.1126/science.258.5088.1647
PubMed Abstract | CrossRef Full Text | Google Scholar
Naghiloo, S., and Claßen-Bockhoff, R. (2017). Udviklingsmæssige ændringer i tid og rum fremmer evolutionær diversificering af blomster: et casestudie i Dipsacoideae. Front. Plant Sci. 8:1665. doi: 10.3389/fpls.2017.01665
PubMed Abstract | CrossRef Full Text | Google Scholar
Nikolov, L. A., Tomlinson, P. B., Manickam, S., Endress, P. K., Kramer, E. M., og Davis, C. C. (2014). Holoparasitiske Rafflesiaceae besidder de mest reducerede endofytter og giver alligevel anledning til verdens største blomster. Ann. Bot. 114, 233-242. doi: 10.1093/aob/mcu114
PubMed Abstract | CrossRef Full Text | Google Scholar
Nodine, M. D., og Bartel, D. P. (2010). MicroRNA’er forhindrer tidlig genekspression og muliggør mønsterdannelse under planteembryogenese. Genes Dev. 24, 2678-2692. doi: 10.1101/gad.1986710
PubMed Abstract | CrossRef Full Text | Google Scholar
Ojeda, D. I., Jaén-Molina, R., Santos-Guerra, A., Caujape-Castells, J., og Cronk, Q. (2017). Tidsmæssige, men ikke rumlige, ændringer i ekspressionsmønstre af kronbladsidentitetsgener er forbundet med tab af papillerede kegleformede celler og skiftet til fuglebestøvning i Macaronesisk Lotus (Leguminosae). Plant Biol. 19, 420-427. doi: 10.1111/plb.12551
PubMed Abstract | CrossRef Full Text | Google Scholar
Park, S. J., Jiang, K., Schatz, M. C., Schatz, M. C., og Lippman, Z. B. (2012). Hastigheden af meristemmodning bestemmer blomsterstandens arkitektur i tomat. Proc. Natl. Acad. Sci. U.S.A. 109, 639-644. doi: 10.1073/pnas.1114963109
PubMed Abstract | CrossRef Full Text | Google Scholar
Peragine, A., Yoshikawa, M., Wu, G., Albrecht, H. L., og Poethig, R. S. (2004). SGS3 og SGS2/SDE1/RDR6 er nødvendige for juvenil udvikling og produktion af transvirkende si RNA’er i Arabidopsis. Genes Dev. 18, 2368-2379. doi: 10.1101/gad.1231804.nism
CrossRef Full Text | Google Scholar
Pham, T., Day, S. M., Glassford, W. J., Williams, T. M., og Rebeiz, M. (2017). Den evolutionære oprindelse af et nyt udtryksmønster gennem et ekstremt heterokronisk skift. Evol. Dev. 19, 43-55. doi: 10.1111/ede.12215
PubMed Abstract | CrossRef Full Text | Google Scholar
Pires, N. D., og Dolan, L. (2012). Morfologisk evolution i landplanter: nye designs med gamle gener. Philos. Trans. R. Soc. B Biol. Sci. 367, 508-518. doi: 10.1098/rstb.2011.0252
PubMed Abstract | CrossRef Full Text | Google Scholar
Poethig, R. S. (2013). Vegetativ faseændring og skudmodning i planter, 1st Edn. New York, NY: Elsevier Inc. doi: 10.1016/B978-0-12-396968-2.00005-1
PubMed Abstract | CrossRef Full Text | Google Scholar
Porras, R., og Muñoz, J. M. (2000). Cleistogamous capitulum hos Centaurea (Asteraceae): heterochron oprindelse. Am. J. Bot. 87, 925-933. doi: 10.2307/2656991
PubMed Abstract | CrossRef Full Text | Google Scholar
Pryer, K. M., og Hearn, D. J. (2009). Udvikling af bladform i marsileaceous bregner: beviser for heterochroni. Evolution 63, 498-513. doi: 10.1111/j.1558-5646.2008.00562.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Ronse de Craene, L. (2018). Forståelse af blomstringsudviklingens rolle i evolutionen af angiospermblomster: afklaringer fra et historisk og fysisk-dynamisk perspektiv. J. Plant Res. 131, 367-393. doi: 10.1007/s10265-018-1021-1
PubMed Abstract | CrossRef Full Text | Google Scholar
Rothwell, G. W., Wyatt, S. E., og Tomescu, A. M. F. (2014). Planteevolution i grænsefladen mellem palæontologi og udviklingsbiologi: et organismcentreret paradigme. Am. J. Bot. 101, 899-913. doi: 10.3732/ajb.1300451
PubMed Abstract | CrossRef Full Text | Google Scholar
Sablowski, R. (2016). Koordinering af plantecellevækst og -deling: kollektiv kontrol eller gensidig aftale? Curr. Opin. Plant Biol. 34, 54-60. doi: 10.1016/j.pbi.2016.09.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Sablowski, R., og Carnier Dornelas, M. (2014). Samspillet mellem cellevækst og cellecyklus i planter. J. Exp. Bot. 65, 2703-2714. doi: 10.1093/jxb/ert354
PubMed Abstract | CrossRef Full Text | Google Scholar
Sharbel, T. F., Voigt, M. L., Corral, J. M., Galla, G., Kumlehn, J., Klukas, C., et al. (2010). Apomictic og seksuelle ægceller af boechera viser heterochroniske globale genekspressionsmønstre. Plant Cell 22, 655-671. doi: 10.1105/tpc.109.072223
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, K. K. (2002). Sekvens heterochroni og udviklingens udvikling. J. Morphol. 252, 82-97. doi: 10.1002/jmor.10014
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, K. K. (2003). Time ‘ s arrow: heterochrony og udvikling af udvikling. Int. J. Dev. Biol. 47, 613-621.
Google Scholar
Stone, S. L., Braybrook, S. A., Paula, S. L., Kwong, L. W., Meuser, J., Pelletier, J., et al. (2008). Arabidopsis leafy cotyledon2 inducerer modningsegenskaber og auxinaktivitet: implikationer for somatisk embryogenese. Proc. Natl. Acad. Sci. U.S.A. 105, 3151-3156. doi: 10.1073/pnas.0712364105
PubMed Abstract | CrossRef Full Text | Google Scholar
Sun, B., Looi, L. S., Guo, S., He, Z., Gan, E. S., Huang, J., et al. (2014). Timing mekanisme afhængig af celledeling påberåbes af Polycomb eviction i plante stamceller. Science 343:1248559. doi: 10.1126/science.1248559
PubMed Abstract | CrossRef Full Text | Google Scholar
Tian, H. Q., Yuan, T., og Russell, S. D. (2005). Forholdet mellem dobbelt befrugtning og cellecyklus i mandlige og kvindelige gameter af tobak. Sex. Plant Reprod. 17, 243-252. doi: 10.1007/s00497-004-0233-9
CrossRef Full Text | Google Scholar
Tomescu, A. M. F., Wyatt, S. E., Hasebe, M., og Rothwell, G. W. (2014). Den tidlige evolution af karplanternes kropsbygning – de manglende mekanismer. Curr. Opin. Plant Biol. 17, 126-136. doi: 10.1016/j.pbi.2013.11.016
PubMed Abstract | CrossRef Full Text | Google Scholar
Tsai, A. Y.-L., og Gazzarrini, S. (2014). Trehalose-6-fosfat og SnRK1-kinaser i planteudvikling og signalering: det nye billede. Front. Plant Sci. 5:119. doi: 10.3389/fpls.2014.00119
PubMed Abstract | CrossRef Full Text | Google Scholar
Vasconcelos, T. N. C., Lucas, E. J., Faria, J. E. Q., og Prenner, G. (2018). Blomstret heterochroni fremmer fleksibilitet af reproduktionsstrategier i den morfologisk homogene slægt Eugenia (Myrtaceae). Ann. Bot. 121, 161-174. doi: 10.1093/aob/mcx142
PubMed Abstract | CrossRef Full Text | Google Scholar
Wahl, V., Ponnu, J., Schlereth, A., Arrivault, S., Langenecker, T., Franke, A., et al. (2013). Regulering af blomstring ved trehalose-6-fosfat-signalering i Arabidopsis thaliana. Science 339, 704-707. doi: 10.1126/science.1230406
PubMed Abstract | CrossRef Full Text | Google Scholar
Walcher-Chevillet, C. L., og Kramer, E. M. (2016). Breaking the mold: forståelse af evolution og udvikling af sideorganer i forskellige plantemodeller. Curr. Opin. Genet. Dev. 39, 79-84. doi: 10.1016/j.gde.2016.06.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Wang, J., Czech, B., og Weigel, D. (2009). miR156-regulerede SPL-transskriptionsfaktorer definerer en endogen blomstringsvej i Arabidopsis thaliana. Cell 138, 738-749. doi: 10.1016/j.cell.2009.06.014
PubMed Abstract | CrossRef Full Text | Google Scholar
Willmann, M. R., Mehalick, A. J., Packer, R. L., og Jenik, P. D. (2011). MicroRNA’er regulerer timingen af embryomodning i Arabidopsis. Plant Physiol. 155, 1871-1884. doi: 10.1104/pp.110.171355
PubMed Abstract | CrossRef Full Text | Google Scholar
Wu, G., Park, M. Y., Conway, S. R., Wang, J., Weigel, D., og Poethig, R. S. (2009). Den sekventielle virkning af miR156 og miR172 regulerer udviklingstidspunktet i Arabidopsis. Cell 138, 750-759. doi: 10.1016/j.cell.2009.06.031
PubMed Abstract | CrossRef Full Text | Google Scholar
Wu, G., og Poethig, R. S. (2006). Tidsmæssig regulering af skududvikling i Arabidopsis thaliana af miR156 og dets mål SPL3. Development 133, 3539-3547. doi: 10.1242/dev.02521
PubMed Abstract | CrossRef Full Text | Google Scholar
Xu, M., Hu, T., Smith, M. R., og Poethig, R. S. (2016). Epigenetisk regulering af vegetativ faseændring i Arabidopsis. Plant Cell 28, 28-41. doi: 10.1105/tpc.15.00854
PubMed Abstract | CrossRef Full Text | Google Scholar
Xu, Y., Guo, C., Zhou, B., Li, C., Wang, H., Zheng, B., et al. (2016). Regulering af vegetativ faseændring af SWI2/SNF2 kromatin remodellering ATPase BRAHMA. Plant Physiol. 172, 2416-2428. doi: 10.1104/pp.16.01588
PubMed Abstract | CrossRef Full Text | Google Scholar
Xu, M., Leichty, A. R., Hu, T., og Poethig, R. S. (2018). H2A.Z fremmer transkriptionen af MIR156A og MIR156C i Arabidopsis ved at lette aflejringen af H3K4me3. Development 145:dev.152868. doi: 10.1242/dev.152868
PubMed Abstract | CrossRef Full Text | Google Scholar
Yamaguchi, N., Huang, J., Xu, Y., Tanoi, K., og Ito, T. (2017). Finjustering af auxinhomeostase styrer overgangen fra vedligeholdelse af blomsterstamceller til gynoeciumdannelse. Nat. Commun. 8:1125. doi: 10.1038/s41467-017-01252-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Yang, L., Xu, M., Koo, Y., He, J., og Poethig, R. S. (2013). Sukker fremmer vegetativ faseændring i Arabidopsis thaliana ved at undertrykke ekspressionen af MIR156A og MIR156C. eLife 2:e00260. doi: 10.7554/eLife.00260
PubMed Abstract | CrossRef Full Text | Google Scholar
Yu, S., Li, C., Zhou, C. M., Zhang, T. Q., Lian, H., Sun, Y., et al. (2013). Sugar is an endogenous cue for juvenile-to-adult phase transition in plants. eLife 2:e00269. doi: 10.7554/eLife.00269
PubMed Abstract | CrossRef Full Text | Google Scholar
Yu, S., Zhang, Y., Horrer, D., Zhang, T., Hao, Y., Feng, Y., et al. (2012). Gibberellin regulerer Arabidopsis blomsterovergangen gennem miR156-målrettede SQUAMOSA-promotorbinding – som transkriptionsfaktorer. Plant Cell 24, 3320-3332. doi: 10.1105/tpc.112.101014
PubMed Abstract | CrossRef Full Text | Google Scholar
PubMed Abstract | CrossRef Full Text | Google Scholar
Leave a Reply