Jižní AmerikaPes křovinný Speothos venaticus

pes křovinný – © Gerald L Zuercher
Amazonian Canids Working Group – Karen DeMatteo a Fernanda Michalski jsou koordinátorkami Amazonian Canids Working Group. Tato pracovní skupina se zaměřuje na čtyři druhy: psa krátkoocasého, lišku krabožravou, psa křovinného a jihoamerické lišky. Amazonské psovité šelmy jsou podobné mnoha šelmám po celém světě, jejichž dlouhodobé přežití je ohroženo řadou přímých i nepřímých hrozeb, včetně ztráty životního prostředí, rozšiřování vodních přehrad, nelegálního lovu kořisti a nemocí způsobených domácími psy.
Projekty
- Proyecto Zorro Pitoco: Využití neinvazivních technik k získání nových poznatků o psech křovinných a koexistující biodiverzitě v Misiones, Argentina
- Monitorování divokých zvířat pomocí kamerových pastí v národním parku Madidi a jeho okolí, Bolívie
- Zkoumání neotropické helmintózy a dalších parazitů v biotopu volně žijících zvířat v pralese Misionera jako potenciálních ohnisek zoonóz
- Vliv klimatických změn na šelmy v Amazonii
Relevantní odkazy
- Flickr:
- 2011 IUCN Red List Assessment – Bush Dog
Reports / Papers
- Residents of the forest – canids from the Amazon
- 2004 Status Survey & Convervation Action Plan – (Přehled stavu psovitých šelem v Amazonii). Jižní Amerika
- Proč se psi z buše tolik liší od ostatních psů
- Vzácní psi z amazonské buše zachycení kamerou
- Záchranářský pes nyní pomáhá zachraňovat jiné (mnohem divočejší) psy
- Úžasný druh: Psi křovináři
Další názvy
Anglický:
Francouzština: Bush Dog, Savannah Dog, Vinegar Dog
Francouzština: Bush Dog, Savannah Dog, Vinegar Dog Chien Des Buissons, Zorro
Španělsky; kastilsky:Cachorro Vinagre, Guanfando, Pero Selvático, Perrito Venadero, Perrito de Monte , Perro De Monte, Perro Grullero , Perro Vinagre , Perro de Agua, Perro de la Selva, Umba, Zorrito Vinagre, Zorro Pitoco , Zorro Vinagre
Něm: Waldhund
italsky: Itticione, Speoto
portugalsky: Cachorrinho, Cachorro-do-mata, Cachorro-do-mato-cotó, Cachorro-do-mato-vinagre, Cachorro-pitoco
Odůvodnění
Přestože je pes křovinář rozšířen, zdá se, že je vzácný v celém svém areálu. Ukázalo se, že je velmi obtížné tento druh ve volné přírodě nalézt, což ztěžuje odhady populačních trendů. Ačkoli lze očekávat, že široce rozšířený druh s potvrzeným výskytem ve fragmentovaných nebo pozměněných biotopech bude mít obecně stabilní populaci, předběžné důkazy z terénu naznačují, že tyto oblasti mohou změnit ekologické nároky psa křovinného, což vede ke zvětšení minimální plochy potřebné k udržení skupiny (a tím ke zvýšení rizika přímého nebo nepřímého vystavení druhu dalším hrozbám). Druh je zde uveden jako téměř ohrožený na základě odhadovaného poklesu o 20-25 % za posledních 12 let (odhadovaná délka generace = 4 roky), což se blíží hranici pro zařazení do kategorie zranitelný podle kritéria A2. Tento úbytek lze přičíst více hrozbám, včetně pokračující ztráty a fragmentace stanovišť, snížení početnosti kořisti v důsledku nelegálního lovu a predace domácími psy a zvýšenému riziku onemocnění způsobených psovitými šelmami.
Informace o geografickém areálu
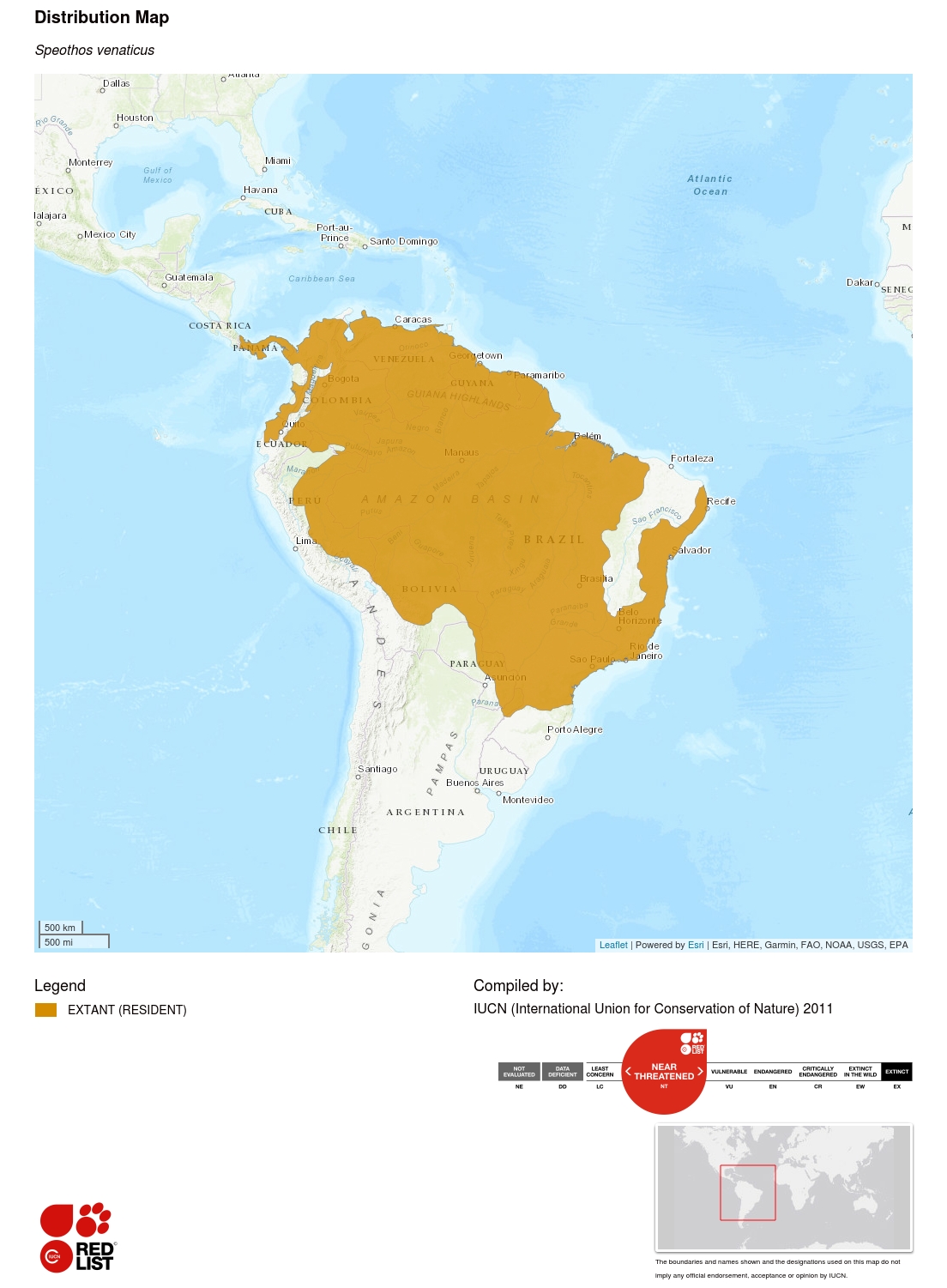
Tento druh se vyskytuje od krajního východu Střední Ameriky a severu Jižní Ameriky, na jih po Paraguay a severovýchodní Argentinu. Izolované subpopulace se mohou stále vyskytovat také v Ekvádoru a Kolumbii západně od And. Jeho historické rozšíření mohlo sahat až na sever Kostariky (Rosa a Nocke 2002), kde druh možná stále přežívá. Současná mapa rozšíření byla vytvořena na základě výsledků rozsáhlého průzkumu biologů zabývajících se šelmami a literatury ohledně výskytu druhu (n = 399 historických lokalit) a také na základě bioklimatického modelu předpovídajícího vhodné oblasti pro tento druh (DeMatteo a Loiselle 2008). Posledně jmenovaní autoři odhadli, že celková předpokládaná plocha vhodná pro psouny křovinné je řádově 14 445 000 km², přičemž 77 % z této plochy se vyskytuje v rámci současného areálu výskytu.
Tendence populace: Klesající
Informace o populaci
Přes velký areál rozšíření a výskyt v různých biotopech se zdá být pes křovinný v celém svém areálu přirozeně vzácný. V nedávném průzkumu, který provedl DeMatteo (2008), většina zemí s výskytem psa křovinného uvedla status druhu jako vzácný nebo neznámý (vzácný: Ekvádor; vzácný nebo neznámý: Argentina, Bolívie, Brazílie, Francouzská Guyana, Paraguay; neznámý: Panama a Venezuela) a pouze dvě země jej uvedly jako běžný (Guyana a Peru). Druh je zřídka zaznamenáván pomocí kamerových pastí; při interpretaci nezjištění je však třeba být opatrný, protože to může ve skutečnosti souviset s tím, že se druh aktivně vyhýbá prvkům spojeným s touto technikou (např, Umístění pastí podél stezek používaných jinými šelmami) nebo předpoklady o pohybu zvířat (např. umístění podél silnic předpokládá, že zvíře půjde podél silnice a nebude ji přecházet).
Demografické údaje o psounech křovinných chybí v celém jejich areálu a odhady populace byly zaznamenány pouze pro několik oblastí: <100 v Misiones Argentina (DeMatteo 2008), >1 000 v Bolívii (DeMatteo 2008), >1 000 na 4 022 km² nebo jeden jedinec/4 km² v Cusco Peru v oblasti řeky Camisea (DeMatteo 2008), 0,04 jedince/km² (B. Beisiegel pers. comm.) v Brazílii, 0,025 jedince/km² v částečně fragmentovaném cerrado v Mato Grosso Brazílie (E.S. Lima, K. E. DeMatteo, R. S. P. Jorge, M. L. S. P. Jorge, J. Dalponte, H. S. Lima a S. Klorfine, pers. obs.), 15,8 jedinců/394 km² nebo 0,04 jedinců/km² v Pantanalu v Brazílii (Lima, Jorge et al. 2009) a 0,001 jedinců/km² ve fragmentované jižní Amazonii Brazílie (Michalski 2010). Vhodný areál o rozloze přibližně 11 000 000 km² (DeMatteo a Loiselle 2008) a průměrná populační hustota přibližně 0,01 jedince/km² by předpovídaly odhadovanou populaci 110 000 jedinců, z nichž přibližně polovina by byla dospělá; vliv fragmentace, stupeň ochrany a variabilita v rámci jejich širokého rozšíření však naznačují, že se jedná o nadhodnocený odhad.
Populační trendy v celém areálu rozšíření psa křovinného jsou uváděny jako neznámé ve dvou zemích (Guyana a Panama), klesající ve dvou (Paraguay a Venezuela), neznámé nebo stabilní v jedné (Francouzská Guyana) a stabilní ve dvou (Ekvádor a Peru, konkrétně Cusco Peru) (DeMatteo 2008). V celém areálu výskytu je vzhledem k rostoucí míře fragmentace biotopů (urbanizace a zemědělství), ohrožení populací kořisti nelegálním pytláctvím a vystavení potenciálně smrtelným chorobám souvisejícím s kandidami velmi nepravděpodobné, že by trend byl stabilní nebo rostoucí.
Informace o biotopu a ekologii
Původní obyvatelé uvádějí, že psoun křovinný je habitatový generalista, a to v kontextu výskytu zpravidla v blízkosti vodních zdrojů, zejména malých toků, kde může být vyšší hustota kořisti (Zuercher et al. 2005; E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima a S. Klorfine pers. obs.; M. Swarner, pers. comm). Křovináři byli pozorováni v nížinných (pod 1 500 m n. m.) lesních biotopech včetně primárních a galerijních lesů (Defler 1986), poloopadavých lesů a sezónně zaplavovaných lesů (Aquino a Puertas 1997). Pozorování byla zaznamenána také z biotopů cerrado v Brazílii (Silveira et al. 1998; Oliveira 2009, C. Brady pers. comm.) a Paraguayi (Zuercher a Villalba 2002) a okrajů pampy (vlhké savany)/riparií (Strahl et al. 1992, Emmons 1998). Nejnovější zprávy zmiňují psouny křovinné ve třech jedinečných biotopech: caatinga, chaco a mangrovové porosty na pobřeží (DeMatteo a Loiselle 2008). V některých případech byli pozorováni i několik kilometrů od lesních biotopů (Silveira et al. 1998). Druh je také příležitostně uváděn ze sekundárních lesů, rančů (M. Swarner pers. comm.), fragmentovaných rančů cerrado (E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima a S. Klorfine pers. obs.; L. Silveira a A. Jácomo pers. comm.), narušených ploch (DeMatteo a Loiselle 2008, Oliveira 2009) a fragmentovaných lesních ploch (Michalski a Peres 2005, Michalski 2010).
Nejsou zjištěny žádné preference výběru stanoviště ani v Pantanalu v Brazílii (Lima, Jorge et al. 2009) ani ve směsi Atlantický les – cerrado v horní Paraná v Paraguayi (Zuercher et al. 2005); byly však nalezeny určité důkazy naznačující preferenci nedotčených savan a lesních stanovišť oproti změněným (orná půda a pastviny) u skupiny psounů křovinných v částečně fragmentovaném cerrado, což naznačuje, že využívání a preference stanovišť se mohou lišit podle regionu a dostupnosti (E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima a S. Klorfine pers. obs.). Zatímco Michalski (2010) uvádí nepřítomnost psounů křovinných ve fragmentovaných oblastech v jižní Amazonii, jiné studie prokázaly silné důkazy o tom, že jejich výskyt je pravděpodobný (DeMatteo a Loiselle 2008; Oliveira 2009; E. S. Lima, K. E. DeMatteo, R. S. P. Jorge, M. L. S. P. Jorge, J. Dalponte, H. S. Lima a S. Klorfine pers. obs.); intenzita využívání těchto oblastí a dopad na ekologické nároky však nejsou známy. Předběžné údaje z terénu naznačují, že s rostoucím stupněm fragmentace biotopů roste i plocha potřebná pro psouny křovinné (E. S. Lima pers. comm.); i když jsou tyto údaje k dispozici pouze pro ekosystém cerrado v Brazílii, předpokládá se, že to platí pro všechny typy biotopů a je to odrazem negativního vztahu mezi hustotou kořisti a fragmentací biotopů. Reanalýza historického rozšíření psouna křovinného (n > 250 lokalit) určila, že přibližně čtvrtina (20 %) byla spojena s fragmentovaným/změněným biotopem a procento změněného biotopu se zvýšilo, když byl odhad rozšířen z plochy potřebné k obživě jednoho jedince na plochu pro hnízdní skupinu (DeMatteo a Loiselle 2008).
Uváděné rozdíly v odhadech velikosti domovských okrsků pravděpodobně souvisejí s celistvostí stanoviště a hustotou kořisti: 150 km² (Horní Paraná, Atlantický les; Beisiegel 1999), ca. 16 km² (Beisiegel a Ades 2004) a 140 km² (Fixed Kernel 95%) pro jednu skupinu psounů křovinných (n = 2-4 jedinci) v cerrado (E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima a S. Klorfine, pers. obs.).
Předběžné terénní údaje podporují studie v zajetí, podle nichž se křovináři vyskytují v rodinných skupinách s mláďaty z jednoho nebo více vrhů (DeMatteo 2008, Michalski 2010, E.S. Lima, K. E. DeMatteo, R. S. P. Jorge, M. L. S. P. Jorge, J. Dalponte, H. S. Lima a S. Klorfine, pers. obs.); pravidelně jsou však pozorováni i samotářští jedinci v různých typech prostředí. Rozmnožování v přírodě je sezónní (DeMatteo 2008). Zdá se, že druh má částečně nomádský způsob pohybu oproti skutečnému teritoriu jako mnohé šelmy (DeMatteo 2008, E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima a S. Klorfine, pers. obs.) a byl zaznamenán žijící v sympatrii s Atelocynus microtis v Peru a Brazílii (Leite Pitman, Beck et al. 2003, Michalski 2010). Odhadovaná délka generace je čtyři roky [(0,5 x délka rozmnožování šest let) + pohlavní dospělost v jednom roce (v zajetí; Porton et al. 1987).
Předběžné terénní údaje podporují neoficiální zprávy o potravě a názor, že křovináři jsou vysoce masožraví (Deutsch 1984, Peres 1991); zdá se však, že existují důkazy o preferenci kořisti v závislosti na geografické oblasti a typu stanoviště, k čemuž pravděpodobně přispívá sezónní proměnlivost. Pro oblast Pantanalu v Brazílii byl hlavní potravou pásovec devítipásý (Dasypus novemcinctus; 94 %), v menším množství se vyskytovali drobní savci, aguti (Dasyprocta azarae) a ptáci; nebylo však zaznamenáno žádné ovoce (n = 17, určeno podle pachu a stop; Lima, Jorge et al. 2009). V případě vnitřního atlantského lesa v Paraguayi tvořili většinu potravy Agoutis a Paca (Cuniculus paca), v menším množství se vyskytovali drobní savci, hlodavci, plazi, bezobratlí a plody Cecropia (n = 11, určeno pomocí mitochondriální DNA; Zuercher et al. 2005). V úspěšném reprodukčním centru v jihovýchodní Brazílii, kde je k dispozici maso a ovoce (banány a papája), se zvířata živí ovocem a masem ve stejném poměru (L. Saboia, pers. comm.). Příležitostně byla zaznamenána predace drůbeže (DeMatteo 2008).
Informace o hrozbách
Existuje několik vážně vnímaných hrozeb, mj: 1) zásah člověka a ztráta nedotčeného biotopu v důsledku velkoplošného zemědělství (např. sóji), přeměny půdy na pastviny a velkoplošných výsadeb monokulturních dřevin (např. stromů), eukalyptus, borovice); 2) snížení početnosti kořisti v důsledku nelegálního pytláctví a predace domácími psy; a 3) zvýšené riziko nákazy smrtelnými nemocemi od domácích psů (blízkost lidské populace a loveckých psů (DeMatteo 2008).
Nemoci související s kanidami jsou hrozbou, která nebyla pro psy křovináře dříve identifikována. Terénní poznatky však potvrzují, že patogeny mohou být přenášeny domácími psy a následky mohou být potenciálně devastující, především kvůli skupinovému způsobu života tohoto druhu (Mann et al. 1980, Steinel et al. 2001, Leite Pitman, Nieto a kol. 2003, Jorge, Morato a kol. 2007, Jorge, Nunes a kol. 2007, DeMatteo 2008, Jorge a kol. 2008, E.S. Lima, K.E. DeMatteo, R.S.P. Jorge, M.L.S.P. Jorge, J. Dalponte, H.S. Lima a S. Klorfine, pers. obs.). E. S. Lima, K. E. DeMatteo, R. S. P. Jorge, M. L. S. P. Jorge, J. Dalponte, H. S. Lima a S. Klorfine (pers. obs.) pozorovali tento účinek, když se generalizované vypadávání srsti, podezřelé jako typ svrabu, postupně rozšířilo ve skupině divokých psů křovinářů a nakonec zahubilo všechny jedince. Kromě svrabu jsou uváděny nemoci parvoviróza a vzteklina, které negativně ovlivňují populace volně žijících psů (Mann et al. 1980, DeMatteo 2008). U tohoto druhu byla zjištěna náchylnost k infekcím Dioctophyma renale i Amphimerus interruptus (muzejní sbírky – Vierira et al. 2008), Lagochilascaris sp. (Volcán a Medrano 1991) a jako hostitel Echinococcus vogeli (Cestoda: Taeniidae) (Rausch a Bernstein 1972). U zvířat chovaných v zajetí byla zjištěna náchylnost k různým chorobám a parazitům, včetně parvovirózy (Janssen et al. 1982), vakcinací vyvolané psinky (McInnes et al. 1992), leishmanie (Lima, Fattori et al. 2009), Spirocerca lupi (Rinas et al. 2009), Toxoplasma gondii (Sedlák a Bartová 2006) a Campylobacter (L. Saboia pers. comm.). U posledně jmenovaného náhle uhynulo 13 z 15 jedinců ve velké rodinné skupině na těžkou hemoragickou enteritidu, když se nakazili kampylobakterem z infikované koaty, kterou byli neúmyslně krmeni (L. Saboia pers. comm.). Tato ztráta více zvířat v krátkém období zdůrazňuje jak citlivost druhu k různým nemocem, tak náchylnost skupinově žijících druhů k patogenům. S rostoucí blízkostí lidských oblastí a zásahy loveckých psů do chráněných území je riziko vystavení nemocem potenciálně vysoké. Expozici nemocem mohou způsobit také zdivočelí nebo polodivocí, neočkovaní, domácí psi, kteří pravidelně loví kořist nezávisle na člověku (K. DeMatteo pers. obs.). DeMatteo (2008) poznamenal, že v Brazílii bylo zaznamenáno více problematických interakcí mezi psy křovináři a domácími psy než s hospodářskými zvířaty, což jen podtrhuje hrozbu nákazy pro tento druh.
Informace o využití a obchodu
Psi křovináři byli zaznamenáni zastřelení v pěti zemích (Argentina, Brazílie, Panama, Peru, Venezuela) a odchycení ve dvou (Panama a Peru) (DeMatteo 2008, F. Michalski nepubl. údaje). V Peru byly tyto události spojeny s tím, že domorodci uváděli, že psouna křovinného snědí, pokud je chycen do pasti, nebo pokud mají vzácnou příležitost ho pak sbírat při tradičním lovu; ve Venezuele však byly hlášené odstřely spojeny s příležitostným sběrem trofejí kvůli jeho vzácnosti (DeMatteo 2008).
Informace o ochranářských opatřeních
Tento druh se vyskytuje v několika chráněných oblastech v celém svém areálu, má však nerovnoměrné rozšíření a vyskytuje se ve velmi nízkých hustotách. Lov je zakázán v Kolumbii (zákon číslo 848:1973), Ekvádoru (zákon číslo 74:1981), Francouzské Guyaně (zákon číslo JO19860625:1986), Panamě (zákon číslo 2-80:1980), Paraguayi (zákon číslo 18796:1975) a Peru (zákon číslo 5056:1970). Lov a obchod s nimi je regulován v Argentině (zákon číslo 22.421:1981), Bolívii (zákon číslo 12301:1975), Brazílii (zákon číslo 5197:191967) a Venezuele (zákon číslo 276:1970). Informace pro Guyanu a Surinam nejsou k dispozici. Bohužel v mnoha částech jeho areálu nejsou dostatečné zdroje pro správu vyhlášených chráněných území a prosazování stávajících zákonů o ochraně volně žijících živočichů.
Zapsán v CITES v příloze I.
Křovináři se vyskytují v zajetí a jsou součástí úspěšného mezinárodního chovného programu (Buck 2009), který zahrnuje Asii (Japonsko), Evropu, Brazílii a Severní Ameriku. Nejsou známy žádné pokusy o reintrodukci.
Odhady populace a demografické údaje o psu křovinném jsou v celém jeho areálu dosud málo známé. To se týká i pochopení sociální dynamiky jednotlivých skupin, zejména pokud jde o rozptyl mláďat a areál využití vzhledem k ostatním skupinám (překrývající se nebo oddělené domovské okrsky). Nejsou jasně známy vazby na stanoviště – druh byl dříve považován za závislý na lesích, ale nyní je stále častěji pozorován na otevřených a fragmentovaných stanovištích; neexistují však žádné údaje o životaschopnosti populace v takových oblastech. Předběžné údaje o potravě potvrzují, že se jedná především o masožravou stravu, je však třeba vyhodnotit sezónní změny a geografické rozdíly v potravě. Je třeba zjistit, jak lze ve volně žijících populacích řídit nebo minimalizovat vliv nemocí, zejména dynamiku přenosu od domácích psů. Je třeba dále vyhodnocovat mezidruhové vztahy se sympatrickými masožravci.
Lokalizace důkazů přítomnosti druhu pomocí standardních průzkumných technik, včetně kamerových pastí a transektových průzkumů, se ukázala jako obtížná (Beisiegel 2009, DeMatteo et al. 2009, Michalski 2010). Použití umělých pachových návnad, které mohou zvýšit přitažlivost k určité lokalitě, bylo u divokých křovinářů neúspěšné (Zuercher et al. 1999). Existuje však několik metodických úprav, které mohou zvýšit účinnost těchto technik u tohoto druhu, včetně úpravy výšky umístění kamery, zvýšení úsilí při odchytu a současného použití přehrávání dlouhých vokálů a konspecifické moči (DeMatteo et al. 2004). Omezené terénní pokusy v částečně fragmentovaném cerrado (2004-2005) s playbacky, močí a pastmi s držením nohou byly neúspěšné (K. DeMatteo nepubl. data). Je však zapotřebí dalších pokusů, aby se zjistilo, jak hustota druhů, variabilita stanovišť (les versus cerrado) a hustota kořisti mění účinnost techniky.
Předběžný a probíhající výzkum využívající kombinaci tří neinvazivních technik (psi s pachovou detekcí, screening DNA trusu, technologie GIS) se u tohoto druhu ukázal jako úspěšný (DeMatteo et al. 2009, nepubl. data) a měl by být rozšířen na další oblasti a stanoviště. Tento soubor technik eliminuje nutnost přilákat druh na konkrétní lokalitu a umožňuje nahlédnout do mnoha ekologických proměnných, včetně využívání biotopu (neporušeného a fragmentovaného), stavu populace, minimální plochy využívání a překrývání/oddělování niky s jinými šelmami.
.
Leave a Reply