Frontiers in Pharmacology
Úvod
Nejčastější typ demence, Alzheimerova choroba (AD), je doprovázen chronickým zhoršováním kognitivních funkcí a představuje 60-70 % případů demence (Fan a Chiu, 2014; Lu a kol., 2018). Podle přehledu z roku 2014 trpí v současné době Alzheimerovou chorobou přibližně 35,6 milionu lidí a její výskyt se s věkem postupně zvyšuje a do roku 2050 postihne celosvětově přibližně 115,4 milionu lidí (Fan a Chiu, 2014). Obecně se uznává, že AD je komplikované neurodegenerativní onemocnění mnohočetných patologií, které souvisí především s degenerací cholinergního přenosu, oxidačním stresem, neurozánětem atd (Coyle et al., 1983; Aucoin et al., 2005; Lin a Beal, 2006; Verri et al., 2012; Heneka et al., 2015; Bassani et al., 2017). Inhibitory acetylcholinesterázy (AChE), jako jsou donepezil, rivastigmin a galanthamin, jsou přednostně předepisovány pro klinickou léčbu AD v časných stadiích (Kwon et al., 2010; Rijpma et al., 2014; Kumar a Singh, 2015). Kromě toho se v adjuvantní terapii AD příslovečně používají také antioxidanty a protizánětlivé látky (Zhou et al., 2016; Demirci et al., 2017). Všechny tyto látky však působí omezeně kvůli postupné ztrátě účinnosti s postupujícím onemocněním a léky jsou také spojeny s mnoha toxickými vedlejšími účinky (Zhou et al., 2016; Jeon et al., 2017; Lu et al., 2018). Konkrétně v současné době není k dispozici žádná uspokojivá léčba, která by AD vyléčila nebo změnila její progresivní průběh.
Harmalin (HAL) a harmin (HAR) jsou hojně se vyskytující farmakologické β-karbolinové alkaloidy Peganum harmala L. s podobnou chemickou strukturou (Li et al., 2017b). Mají významnou inhibiční aktivitu AChE, monoaminooxidázy (MAO) a myeloperoxidázy (MPO), antioxidační, protizánětlivé, protinádorové a antihypertenzní účinky a mohou také ovlivňovat obsah různých neurotransmiterů, pak mohou vést k biochemickým, fyziologickým a behaviorálním změnám u zvířat a lidí (Mahmoudian et al., 2002; Frison et al., 2008; Jiménez et al., 2008; Herraiz et al., 2010; Louis et al., 2010; Bensalem et al., 2014; Wang et al., 2015; Li et al., 2017a). Podle našich předchozích studií vykazovaly HAL a HAR podobnou inhibici AChE ve srovnání s galanthaminem in vitro (Zhao et al., 2013). Navíc celkové alkaloidy (28 mg/kg) P. harmala obsahující především HAL a HAR vykazovaly zlepšující účinek na deficity učení a paměti vyvolané skopolaminem a 30% ethanolem (He et al., 2015b). Následně studie He et al. (2015a) dále potvrdila, že HAR (20 mg/kg) může zlepšit poruchy paměti myší vyvolané skopolaminem posílením cholinergních funkcí prostřednictvím inhibice AChE. Nicméně nejnovější výzkum ukázal, že HAR je nejen substrátem izoformy 2 proteinu asociovaného s více léky (MRP2), ale má také slabou metabolickou stabilitu a nakonec vede k podstatně nižší biologické dostupnosti než HAL u různých zvířat (Li et al., 2017b). Spekuluje se tedy, že účinná dávka HAL proti AD může být mnohem nižší než HAR. Proto by měla být provedena systematická srovnávací studie antiamnestických účinků a mechanismů obou analogů s cílem hledat vhodnější kandidátní sloučeninu pro terapii AD.
V současné studii byly porovnány modulační účinky HAL a HAR na poruchy paměti vyvolané skopolaminem pomocí behaviorálního hodnocení v Morrisově vodním bludišti (MWM). MWM se hojně používá při studiu neurobiologie a neurofarmakologie prostorového učení a paměti a který hraje důležitou roli při ověřování modelů hlodavců pro neurokognitivní poruchy, jako je AD (Bromley-Brits et al., 2011). Dále byly komplexně provedeny biochemické testy, western blotting a také imunofluorescenční analýza, aby se objasnily možné účinky HAL a HAR na některé základní mechanismy podílející se na progresi AD. Výsledky této studie budou přínosné pro hodnocení antiamerických účinků β-karbolinových alkaloidů v dalším výzkumu a vývoji a mohly by poskytnout cenné informace pro klinickou léčbu AD.
Materiály a metody
Materiály
Harmalin, HAR, L-tryptofan (L-Trp), 5-hydroxytryptamin (5-HT), 5-hydroxyindol-3-octová kyselina (5-HIAA), acetylcholinchlorid (ACh), cholinchlorid (Ch), γ-aminomáselná kyselina (γ-GABA), monosodná sůl kyseliny L-glutamové (L-Glu), L-fenylalanin (L-Phe), L-tyrosin (L-Tyr), theofylin a všechny ostatní použité chemikálie byly zakoupeny od společnosti Sigma-Aldrich, Co., Ltd. (St. Louis, MO, Spojené státy). Skopolamin hydrobromid a donepezil hydrochlorid monohydrát byly zakoupeny od společnosti TCI (Shanghai) Development, Co, Ltd., v USA. (Šanghaj, Čína). Karboxymethylcelulosa sodná (CMC-Na) a NaCl byly získány od společnosti Meilunbio® Biotech, Co., Ltd. (Čína). (Dalian, Čína). Složené barvivo oxid titaničitý bylo zakoupeno od Shanghai Dyestuffs Research Institute, Co., Ltd. (Čína). (Šanghaj, Čína). Souprava pro kvantifikaci bílkovin BCA, lyzační pufr RIPA, hovězí sérový albumin (BSA), PBST (10X), 30% akrylátový amid, 10% SDS, glycin, pufr pro nakládání vzorků (4X) a TEMED byly zakoupeny od společnosti YEASEN Biotechnology, Co. (Šanghaj, Čína). Inhibitor fosfolipázy a inhibitor proteázy byly zakoupeny od společnosti Roche Applied Science (Foster City, CA, Spojené státy). Membrána PVDF a chemiluminiscenční substrát HRP ImmobilonTM Western byly zakoupeny od společnosti Millipore (Billerica, MA, Spojené státy). Králičí anti-AChE, králičí anti-cholin acetyltransferáza (ChAT), králičí anti-MPO a anti-glyceraldehyd 3-fosfát dehydrogenáza (GAPDH), protilátky proti králičímu IgG konjugované s HRP a marker byly zakoupeny od společnosti Abcam Technology (Cambridge, MA, Spojené státy). Acetonitril, metanol a kyselina mravenčí třídy HPLC byly zakoupeny od společnosti Fisher Scientific, Co. (Santa Clara, CA, Spojené státy). Deionizovaná voda (>18 mΩ) byla přečištěna pomocí systému Milli-Q Academic (Millipore, Corp., Billerica, MA, Spojené státy). Všechny ostatní chemikálie byly analytické kvality.
Zvířata
Sto osm myších samců C57BL/6 (ve věku 10 týdnů) bylo získáno z Centra pro hodnocení bezpečnosti léčiv a výzkum Šanghajské univerzity tradiční čínské medicíny. Myši byly chovány v dobře osvětlené klimatizované místnosti (25 ± 1 °C) za standardních podmínek prostředí (relativní vlhkost: 60-65 %, 12hodinové cykly světlo-tma: světlo svítí od 7:00 do 19:00) a měly volný přístup ke krmivu pro hlodavce a vodovodní vodě. Všechny postupy při použití zvířat byly v souladu s předpisy pro pokusy na zvířatech vydanými Státním výborem pro vědu a technologii Číny dne 14. listopadu 1988 a schválenými Etickou komisí pro pokusná zvířata Šanghajské univerzity tradiční čínské medicíny (č. 1). SUTCM-2011-1107; datum schválení: 10. listopadu 2011).
Podávání léčiv
Na základě rozdílu v biologické dostupnosti HAL a HAR byly stanoveny dávky 2, 5 a 10 mg/kg pro HAL a 10, 20 a 30 mg/kg pro HAR (Zhang, 2013; Shi et al., 2014). Sto osm myší bylo náhodně rozděleno do devíti skupin (12 myší na skupinu), a to do kontrolní skupiny (vehikulum, 0,5% CMC-Na), skupiny se skopolaminem (1 mg/kg), skupiny s donepezilem (pozitivní kontrola, 5 mg/kg), skupiny s HAL (2, 5 a 10 mg/kg, tj. nízké: L, střední: M a vysoké: H dávky) a skupiny s HAR (10, 20 a 30 mg/kg, tj. L, M a H dávky). Jak je znázorněno na obrázku 1, před testem MWM byl kontrolní skupině a skupině se skopolaminem perorálně podáván 0,5% roztok CMC-Na a skupinám HAL (dávky L, M a H), HAR (dávky L, M a H) a donepezil (5 mg/kg) byl podáván perorálně po dobu sedmi po sobě následujících dnů. Od 8. do 16. dne byl všem skupinám s výjimkou kontrolní skupiny, která dostávala 30 minut po různých ošetřeních normální fyziologický roztok, intraperitoneálně (i.p.) aplikován skopolamin (1 mg/kg). Následně byly denně prováděny behaviorální testy 30 min po i.p. injekci skopolaminu nebo normálního fyziologického roztoku.
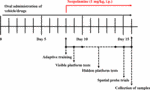
Obrázek 1. Časový rozvrh experimentálního a léčebného designu myší.
Hodnocení chování v Morrisově vodním bludišti
K hodnocení účinků HAL a HAR na prostorové učení a paměť myší byl použit test MWM, který již dříve popsal Richard Morris v roce 1982 s drobnými úpravami (Morris et al., 1982; He et al., 2015a; Singh et al., 2016).
Zařízení MWM se skládalo z bílé kruhové vodní nádrže (průměr: 120 cm a výška: 50 cm) obklopené různými vizuálními značkami (hvězda, čtverec, obdélník a kruh atd.) na sloupcích v pevných pozicích během celého experimentu. Nádrž byla vybělena přidáním netoxického barviva, sloučeniny oxidu titaničitého, při teplotě 22 ± 2 °C, takže plošina byla pod vodní hladinou neviditelná. Nádrž byla prakticky rozdělena na čtyři stejné kvadranty, včetně jihovýchodního, severovýchodního, jihozápadního a severozápadního. Bílá plošina (průměr: 10 cm a výška: 25 cm) byla soustředěna v jihozápadním kvadrantu (Jeon et al., 2017; Kouémou et al., 2017; Lu et al., 2018). Celý experimentální postup se skládal z adaptivního tréninku (1 den, jednou denně), testu viditelné plošiny (1 den, čtyři pokusy denně), testů skryté plošiny (5 dní, čtyři pokusy denně) a testu prostorové sondy (24 h po posledním testu skryté plošiny, jednou denně).
Pro adaptivní trénink (1 den, jednou denně), první den testu MWM (8. den léčby léky) byla každá myš umístěna do vodní nádrže, aby plavala 60 s a absolvovala aklimatizaci. Druhý den (9. den léčby léky) byla za účelem vyšetření vizuální, senzorické a motorické schopnosti myší umístěna viditelná plošina 1 cm nad vodní hladinu a každá myš měla za úkol najít plošinu za 60 s. Test (1 den, čtyři pokusy denně) byl proveden před pokusem se skrytou plošinou za účelem prověření kvalifikovaných myší. Následně byla plošina ponořena 1 cm pod vodní hladinu při testech skryté plošiny (5 dní, čtyři pokusy denně, 10.-14. den léčby léky). Plavecká aktivita byla zaznamenávána videokamerou nad hlavou a analyzována prostřednictvím počítačového systému pro sledování a analýzu obrazu (RD1101-MWM-G, Shanghai Mobile Datum Information Technology Company, Šanghaj, Čína). Myši byly vypuštěny do nádrže s hlavou směřující ke stěně nádrže postupně z jednoho ze čtyř kvadrantů (severovýchod, sever, východ a jihovýchod) a bylo jim umožněno najít plošinu za 60 s. Pokud se myši nepodařilo plošinu najít, měla být jemně navedena konkrétním experimentátorem a měla být ponechána 15 s na plošině. Po každém pokusu byla každá myš odvedena do své klece a nechala se vysušit pod topením. U každého pokusu byla zaznamenána latence úniku a rychlost úniku. Při pokusu s prostorovou sondou (15. den léčby léky) byla plošina z nádrže odstraněna. Každá myš byla umístěna do vodní nádrže z kvadrantu (pro okamžité umístění severozápadní) opačného než předchozí umístění plošiny (cílový kvadrant), aby absolvovala 60sekundový test retence paměti (Lee et al., 2016; Liu et al., 2017; Malik et al., 2017). Zaznamenával se čas v cílovém kvadrantu a počet křížení.
Sběr mozkových tkání
16. den po léčbě léky byly všechny myši anestetizovány a při oftalmektomii byla odebrána plazma a poté byly po dekapitaci odebrány mozkové tkáně. V každé skupině byl mozek dílčího souboru zvířat použit pro imunofluorescenční analýzu a ostatní pro biochemickou analýzu a analýzu Western blotting. Po obětování byly mozkové tkáně okamžitě excidovány, opláchnuty fyziologickým roztokem, filtračním papírem zbaveny povrchové vlhkosti vody a zváženy. Kůra a hipokampus každého mozku byly na ledu rozděleny na dvě části (levou a pravou), resp. Část z nich byla fixována 4% paraformaldehydem pro imunofluorescenční analýzu a ostatní byly ihned zmraženy tekutým dusíkem a poté uloženy při -80 °C až do analýzy.
Biochemický test
Kortex a hipokampus byly homogenizovány v ledové vodě o teplotě 0,5 °C a následně uloženy do ledové vody.1 M fosfátovém pufru (pH 7,4) a 10% (w/v) homogenáty byly centrifugovány při 4 °C při 12 000 otáčkách za minutu po dobu 10 minut a supernatanty byly odděleny a poté uloženy při -80 °C pro biochemické hodnocení (Malik et al., 2017). Byly stanoveny hladiny AChE, ChAT, superoxiddismutázy (SOD), glutathionperoxidázy (GSH-px), maleinového diadehydu (MDA), MPO, tumor nekrotizujícího faktoru α (TNF-α), interleukinu 1β (IL-1β), interleukinu 6 (IL-6), interleukinu 10 (IL-10), a oxidu dusnatého (NO) v homogenátech mozkové kůry a/nebo hipokampu byly stanoveny metodou ELISA pomocí komerčně dostupných testovacích souprav (Jiancheng, Nanjing, Čína) podle protokolů výrobce. Zejména hladiny výše uvedených biochemických faktorů v mozkové kůře a hipokampu byly normalizovány pomocí koncentrace bílkovin každého vzorku. Testy byly provedeny ve dvou opakováních.
Western Blotting Analysis
Mozková kůra a hipokampus byly homogenizovány v ledově chladném 50 mM pufru Tris-HCl (pH 7,4) obsahujícím inhibitor fosfatázy a inhibitor proteázy. Po centrifugaci (12 000 ot./min) po dobu 10 min při 4 °C byl stanoven celkový obsah bílkovin pomocí soupravy BCA protein assay. Vzorky proteinů byly smíchány se čtvrtinovým objemem nakládacího pufru a poté zahřívány 5 min při 100 °C. 20 μg proteinu z každého vzorku bylo elektroforézováno pomocí 8% SDS-PAGE a přeneseno na PVDF membrány. Poté byla membrána 1 h blokována 5% odtučněným mlékem při pokojové teplotě a inkubována s anti-GAPDH (1:5000), anti-AChE (1:1000), anti-ChAT (1:1000) a anti-MPO (1:1000) při 4 °C přes noc. Následně byly membrány důkladně opláchnuty PBST a poté inkubovány s HRP-konjugovanou anti-králičí (1:5000) sekundární protilátkou po dobu 2 h při pokojové teplotě. Po úplném promytí PBST byly proteinové pásy vizualizovány pomocí soupravy ECL prime.
Imnofluorescenční analýza
Tvorba myeloperoxidázy byla vizualizována pomocí imunofluorescenční mikroskopie, jak bylo popsáno dříve s drobnými změnami (Gray et al., 2008). Po fixaci ve 4% paraformaldehydu byla kůra rozřezána na 5 μm silné sagitální řezy, které byly deparafinovány a zpracovány pro imunofluorescenční analýzu. Mozkové řezy byly umístěny do roztoku kyseliny citronové (pH 6,0) pro opravu antigenu v mikrovlnné troubě. Následně bylo provedeno zhášení autofluorescence po dobu 5 min. Po zablokování v 2% BSA byly řezy sondovány anti-MPO (1:100) přes noc při 4 °C. Poté byly řezy třikrát promyty PBS (pH 7,4) a inkubovány se sekundární protilátkou proti králíkovi konjugovanou s HRP (1:300) po dobu 30 min při pokojové teplotě. Poté byl použit 4′,6-diamidino-2-fenylindol (DAPI) pro protibarvení DNA a řezy mozkem byly pozorovány pod fluorescenčním mikroskopem (Nikon ECLIPSE C1, Japonsko) a snímky byly pořízeny odpovídajícím způsobem. Fluorescence DAPI nebo MPO zde byla měřena při excitační vlnové délce 330-380 nebo 510-560 nm a emisní vlnové délce 420 nebo 590 nm.
Vliv HAL a HAR na neurotransmitery
Extrakční postup
K přípravě homogenátů mozkové kůry a vzorků plazmy byla použita rychlá, spolehlivá a pohodlná precipitační metoda. Do centrifugační zkumavky o objemu 1,5 ml bylo přidáno 100 μl vzorku s 200 μl ledově chladného acetonitrilu obsahujícího 60 ng/ml IS a 1 min. se míchalo vortexem. Poté byla směs odstřeďována při 12 000 otáčkách za minutu po dobu 10 min (4 °C). Alikvot 5 μl supernatantu z každého vzorku byl vstříknut do ultraúčinné kapalinové chromatografie kombinované s kvadrupólovou hmotnostní spektrometrií s ionizací elektrosprejem (ESI) (UPLC-ESI-MS/MS) pro kvantitativní analýzu.
UPLC-ESI-MS/MS analýza
Koncentrace různých neurotransmiterů byly simultánně kvantifikovány pomocí systému SHIMADZU LC-30AD UPLC (Shimadzu, Kyoto, Japonsko) připojeného k hmotnostnímu spektrometru AB Sciex QTRAP® 6500 s trojitým kvadrupólem (SCIEX, Spojené státy) vybavenému zdrojem ESI s použitím režimu detekce pozitivních iontů pro monitorování vícenásobných reakcí. Chromatografická separace byla provedena pomocí kolony ZIC-cHILIC (150 mm × 2,1 mm, 3 μm, Merck-Sequant, Německo) s ochrannou kolonou SeQuant ZIC-cHILIC (20 mm × 2,1 mm, 5 μm, Merck-Sequant, Německo). Mobilní fáze, acetonitril-voda (65:35, v/v) obsahující 0,1 % kyseliny mravenčí, byla dodávána při průtoku 0,2 ml/min. Z důvodu ochrany hmotnostního spektrometru před kontaminací byl průtok během první minuty po nástřiku odkloněn do odpadu. Všechny ostatní instrumentální parametry byly nastaveny podle naší předchozí studie (Jiang, 2016). Metoda UPLC-ESI-MS/MS byla dobře ověřena a byla úspěšně použita ke stanovení koncentrací neurotransmiterů v homogenátech mozkové kůry a plazmě (Jiang, 2016).
Statistická analýza
Statistické vyhodnocení bylo provedeno pomocí softwaru SPSS verze 18.0 a data byla vyjádřena jako průměr ± SD. Prezentované údaje o latenci útěku a délce cesty v testu MWM byly analyzovány pomocí opakovaných měření dvoucestné ANOVA. Ostatní behaviorální údaje a změny biomarkerů in vitro byly testovány pomocí jednocestné ANOVA pro vícenásobná srovnání. Významné výsledky byly označeny podle konvenčních kritických P-hodnot: #P < 0,05; ##P < 0,01; ###P < 0,001, oproti kontrolní skupině. ∗P < 0,05; ∗∗P < 0,01; ∗∗∗P < 0,001, oproti skupině vyvolané skopolaminem.
Výsledky
HAL a HAR zmírnily kognitivní dysfunkci myší léčených skopolaminem v MWM
Pro zjištění účinků HAL (dávky L, M a H) a HAR (dávky L, M a H) na prostorové učení a paměť byla u myší s poruchou paměti vyvolanou skopolaminem provedena úloha MWM. Dvoucestná analýza opakovaných měření se smíšeným modelem ukázala, že latence k dosažení plošiny se významně lišila mezi experimentálními skupinami myší (F(8,396) = 26,011, P < 0,001) a dny akvizičního tréninku (F(4,396) = 15,274, P < 0,001). Mezi skupinami a dny nebyla zjištěna žádná významná interakce (F(32,396) = 1,132, P > 0,05), což naznačuje, že rozdíly mezi skupinami byly závislé na léčbě. Jak je uvedeno na obrázku 2, při testování skryté plošiny byly latence útěku a délka cesty ve skupině se skopolaminem významně delší než u kontrolní skupiny ve čtvrtém dni (obrázek 2A, F(8,99) = 3,135, P < 0.01; Obrázek 2B, F(8,99) = 4,074, P < 0,01) a pátý den (Obrázek 2A, F(8,99) = 4,164, P < 0,01; Obrázek 2B, F(8,99) = 4,548, P < 0,01), což svědčí o úspěšně zkonstruovaném modelu poškození paměti u myší. Kognitivní deficity vyvolané skopolaminem však byly zvráceny jak HAL, tak HAR. Ve srovnání se skupinou vyvolanou skopolaminem se u skupin léčených HAL (dávky L, M a H) a HAR (dávky M a H) účinně snížila latence útěku a délka cesty na čtvrté (obr. 2A, F(8,99) = 4,975, P < 0,01; F(8,99) = 2.).816, P < 0,05; F(8,99) = 2,795, P < 0,05; obrázek 2B, F(8,99) = 3,836, P < 0,01; F(8,99) = 3.231, P < 0,01; F(8,99) = 2,658, P < 0,05; obrázek 2C, F(8,99) = 4,673, P < 0,01; F(8,99) = 4,881, P < 0.01; obrázek 2D, F(8,99) = 3,817, P < 0,01; F(8,99) = 3,129, P < 0,01) a pátý den (obrázek 2A, F(8,99) = 12,637, P < 0,001; F(8,99) = 2,802, P < 0,05; F(8,99) = 2,772, P < 0,05; obrázek 2B, F(8,99) = 4.014, P < 0,01; F(8,99) = 2,722, P < 0,05; F(8,99) = 2,531, P < 0,05; obrázek 2C, F(8,99) = 4,574, P < 0.01; F(8,99) = 4,632, P < 0,01; obrázek 2D, F(8,99) = 2,547, P < 0,05; F(8,99) = 3,905, P < 0,01). Ačkoli při léčbě nízkou dávkou HAR v dávce 10 mg/kg (dávkovací skupina L) nedošlo k významnému snížení latence útěku a délky dráhy po celou dobu tréninku na čtvrté (obrázek 2C, F(8,99) = 1.182, P > 0,05; obrázek 2D, F(8,99) = 0,494, P > 0,05) a pátý den (obrázek 2C, F(8,99) = 0,873, P > 0,05; obrázek 2D, F(8,99) = 0,201, P > 0,05). Z obrázku 2 je také patrné, že úniková latence a délka dráhy u skupiny léčené donepezilem (5 mg/kg) se výrazně zkrátily ve srovnání se skupinou léčenou skopolaminem na čtvrtém (obrázek 2A, F(8,99) = 2,668, P < 0.05; Obrázek 2B, F(8,99) = 2,643, P < 0,05) a pátý den (Obrázek 2A, F(8,99) = 2,780, P < 0,05; Obrázek 2B, F(8,99) = 2,897, P < 0,05) během třetího tréninkového pokusného sezení.
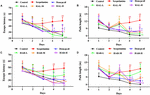
Obrázek 2. Účinky přípravků HAL (2, 5 a 10 mg/kg, konkrétně nízké: L, střední: M a vysoké: H dávky) a HAR (10, 20 a 30 mg/kg, konkrétně L, M a H dávky) na poruchy paměti vyvolané skopolaminem u myší. HAL (dávky L, M a H), HAR (dávky L, M a H) a donepezil (5 mg/kg) byly podávány perorálně 7 dní před tréninkem na Morrisovo vodní bludiště (MWM) a pokračovaly během testování. Poruchy paměti byly vyvolány skopolaminem (1 mg/kg, i.p.). Úniková latence (A) a délka dráhy (B) skupin léčených HAL (dávky L, M a H) v testech se skrytou plošinou po dobu pěti po sobě následujících dnů a úniková latence (C) a délka dráhy (D) skupin léčených HAR (dávky L, M a H) v testech se skrytou plošinou po dobu pěti po sobě následujících dnů. Chybová úsečka (SD) pro datové body na tomto obrázku nebyla uvedena, aby bylo možné jasně rozlišit rozdíly mezi ošetřeními. N = 12/skupina. ##P < 0,01, oproti kontrolní skupině. ∗P < 0,05; ∗∗P < 0,01; ∗∗∗P < 0,001, vs. skupina se skopolaminem.
Den po testování skryté plošiny byl proveden pokus s prostorovou sondou odebráním plošiny za účelem vyhodnocení prostorové paměti všech myší. Jak je znázorněno na obrázku 3, frekvence procházení se ve skupině vyvolané skopolaminem pozoruhodně snížila (F(8,81) = 5,498, P < 0,05), což ilustruje, že 1 mg/kg skopolaminu poškodilo tvorbu prostorové paměti u myší. Zejména kromě skupiny léčené HAR (dávka L) (F(8,81) = 0,494, P > 0,05) ostatní skupiny léčené HAL (dávky L, M a H) a HAR (dávky M a H) dramaticky zvýšily frekvenci míjení (F(8,81) = 11,667, P < 0,05).01; F(8,81) = 9,072, P < 0,01; F(8,81) = 4,507, P < 0,05; F(8,81) = 7,063, P < 0,05; F(8,81) = 8,583, P < 0,01), přičemž nárůst u skupiny s dávkováním HAL-L byl ve srovnání s ostatními skupinami výraznější (obr. 3). Kromě toho se u skupiny léčené donepezilem (5 mg/kg) také významně zvýšila frekvence míjení ve srovnání s myší, které byly léčeny skopolaminem (obr. 3, F(8,81) = 5,345, P < 0,05). Všechny tyto výsledky naznačují, že HAL (dávky L, M a H) a HAR (dávky M a H) mohou zlepšit schopnost prostorové paměti u myší s kognitivní dysfunkcí vyvolanou skopolaminem a HAL vykazuje silnější účinky než HAR na prostorové učení a paměť u MWM.
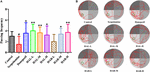
Obrázek 3. Účinky HAL (2, 5 a 10 mg/kg, konkrétně nízké: L, střední: M a vysoké: H dávky) a HAR (10, 20 a 30 mg/kg, konkrétně L, M a H dávky) na poruchy paměti vyvolané skopolaminem u myší. Frekvence průchodu myší místem s plošinou (A) a plavecké dráhy myší ve vodní nádrži (B). Zelený kruh označuje umístění skryté plošiny, zatímco červené křivky představují stopy pohybu myší. N = 10 skupin. #P < 0,05 oproti kontrolní skupině. ∗P < 0,05; ∗∗P < 0,01; vs. skupina se skopolaminem.
Vliv HAL a HAR na skopolaminem indukovaný cholinergní systém, oxidační stres a zánět
Pro další objasnění možných mechanismů působení HAL a HAR na zlepšení paměti vyvolané skopolaminem byly zkoumány hladiny různých biochemických faktorů a exprese proteinů souvisejících s cholinergním systémem, oxidačním stresem a zánětem. Na obrázcích 4, 5 jsou uvedeny změny různých biochemických faktorů v mozkové kůře nebo hipokampu a na obrázcích 6-8 jsou uvedeny účinky HAL a HAR na expresi proteinů v mozkové kůře myší.
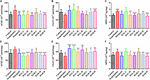
OBRÁZEK 4. Vliv HAL a HAR na expresi proteinů v mozkové kůře myší. Účinky HAL (2, 5 a 10 mg/kg, a to nízký: L, střední: M a vysoký: H) a HAR (10, 20 a 30 mg/kg, konkrétně dávky L, M a H) na aktivity acetylcholinesterázy (AChE), cholinacetyltransferázy (ChAT) a myeloperoxidázy (MPO) v mozkové kůře (A-C) a hipokampu (D-F) myší s kognitivními poruchami vyvolanými skopolaminem. Údaje byly vyjádřeny jako průměr ± SD (n = 10). #P < 0,05; ##P < 0,01; ###P < 0,001, oproti kontrolní skupině. ∗P < 0,05; ∗∗P < 0,01; ∗∗∗P < 0,001, oproti skupině vyvolané skopolaminem.
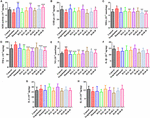
OBRÁZEK 5. Účinky přípravku HAL (2, 5 a 10 mg/kg, a to nízký: L, střední: M a vysoký: H) a HAR (10, 20 a 30 mg/kg, konkrétně dávky L, M a H) na aktivitu nebo produkci superoxiddismutázy (SOD) (A), glutathionperoxidázy (GSH-px) (B), maleinového diadehydu (MDA) (C), tumor nekrotizujícího faktoru α (TNF-α) (D) a oxidu dusnatého (NO) (E) v mozkové kůře myší s kognitivními poruchami vyvolanými skopolaminem. Údaje byly vyjádřeny jako průměr ± SD (n = 10). #P < 0,05; ##P < 0,01; ###P < 0,001, oproti kontrolní skupině. Interleukin 1β (IL-1β) (F), interleukin 6 (IL-6) (G) a interleukin 10 (IL-10) (H). ∗P < 0,05; ∗∗P < 0,01; ∗∗∗P < 0,001, oproti skupině navozené skopolaminem.
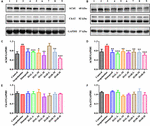
OBRÁZEK 6. Vlivem skopolaminu vzniká v kontrolní skupině zvýšená hladina IL-1β. Účinky HAL (2, 5 a 10 mg/kg, konkrétně nízké: L, střední: M a vysoké: H dávky) a HAR (10, 20 a 30 mg/kg, konkrétně L, M a H dávky) na hladiny exprese proteinů acetylcholinesterázy (AChE), a cholin acetyltransferázy (ChAT) v mozkové kůře (A,C,E) a hipokampu (B,D,F) myší s kognitivními poruchami vyvolanými skopolaminem. Údaje byly vyjádřeny jako průměr ± SD (n = 3). #P < 0,05; ##P < 0,01, oproti kontrolní skupině. ∗P < 0,05; ∗∗P < 0,01; ∗∗∗P < 0,001, oproti skupině vyvolané skopolaminem.

OBRÁZEK 7. V případě, že se jedná o skupinu se skopolaminem, je možné, že se jedná o skupinu se skopolaminem. Účinky HAL (2, 5 a 10 mg/kg, konkrétně nízké: L, střední: M a vysoké: H dávky) a HAR (10, 20 a 30 mg/kg, konkrétně L, M a H dávky) na hladiny exprese proteinu myeloperoxidázy (MPO) v mozkové kůře (A,C) a hipokampu (B,D) myší s kognitivními poruchami vyvolanými skopolaminem. Údaje byly vyjádřeny jako průměr ± SD (n = 3). #P < 0,05; ##P < 0,01, oproti kontrolní skupině. ∗P < 0,05; ∗∗P < 0,01; ∗∗∗P < 0,001, oproti skupině vyvolané skopolaminem.

OBRÁZEK 8. Účinky HAL (2, 5 a 10 mg/kg, konkrétně nízké: L, střední: M a vysoké: H dávky) a HAR (10, 20 a 30 mg/kg, konkrétně L, M a H dávky) na tvorbu myeloperoxidázy (MPO) v mozkové kůře myší s kognitivními poruchami vyvolanými skopolaminem vizualizované pomocí imunofluorescenční mikroskopie.
Vliv HAL a HAR na cholinergní deficity
Pro objasnění účinků HAL a HAR na mozkovou cholinergní funkci byly hodnoceny změny aktivit a hladin proteinů AChE a ChAT v korových a hipokampálních tkáních všech myší. Jak ukazují obrázky 4A,D, léčba skopolaminem v dávce 1 mg/kg významně zvýšila aktivitu AChE v kůře a hipokampu (F(8,81) = 10,553, P < 0,01; F(8,81) = 6,177, P < 0,05). Kromě skupiny s dávkováním HAR-L v kůře ostatní skupiny léčené HAL (dávky L, M a H) a HAR (dávky M a H) výrazně snížily aktivitu AChE (obr. 4A, F(8,81) = 10,837, P < 0,01; F(8,81) = 8,621, P < 0,05; F(8,81) = 15,177, P < 0,05).01; F(8,81) = 8,615, P < 0,05; F(8,81) = 16,330, P < 0,001; obrázek 4D, F(8,81) = 46,593, P < 0,001; F(8,81) = 34,179, P < 0.001; F(8,81) = 7,968, P < 0,05; F(8,81) = 4,769, P < 0,05; F(8,81) = 49,802, P < 0,001; F(8,81) = 46,665, P < 0,001). Stejně tak skupina léčená donepezilem zjevně snížila aktivitu AChE ve srovnání s hladinami ve skupině léčené skopolaminem (obr. 4A, F(8,81) = 13,138, P < 0,01; obr. 4D, F(8,81) = 7,400, P < 0,05). Skupina se skopolaminem navíc vykazovala významné snížení aktivity ChAT v kůře i hipokampu (obr. 4B, F(8,81) = 5,481, P < 0,05; obr. 4E, F(8,81) = 4,874, P < 0,05). Nicméně podávání HAL (dávky L a M) a HAR (dávky M a H) znatelně zvýšilo aktivitu ChAT v kůře (obr. 4B, F(8,81) = 7,956, P < 0,05; F(8,81) = 8,390, P < 0,05; F(8,81) = 7.835, P < 0,05; F(8,81) = 51,927, P < 0,001) a hladiny ChAT ve všech skupinách léčených HAL a HAR v hipokampu byly významně vyšší než ve skupině léčené skopolaminem (obr. 4E, F(8,81) = 18. Obr. 4E.139, P < 0,001; F(8,81) = 9,665, P < 0,01; F(8,81) = 14,113, P < 0,01; F(8,81) = 5,178, P < 0,05; F(8,81) = 7,599, P < 0.05; F(8,81) = 11,953, P < 0,01) a vliv skupiny s dávkováním HAL-L na aktivitu ChAT byl významnější ve srovnání se skupinami s podáváním HAR.
Jak je znázorněno na obrázku 6, ve srovnání s kontrolní skupinou byly hladiny exprese proteinu AChE v mozkové kůře a hipokampu skupiny vyvolané skopolaminem výrazně zvýšeny (obrázek 6C, F(8,18) = 36,632, P < 0,01; obrázek 6D, F(8,18) = 14,823, P < 0,05), zatímco hladiny proteinu ChAT se téměř nezměnily (obrázky 6E,F). Po léčbě HAL (dávky L, M a H), HAR (dávky M a H) a donepezilem, s výjimkou HAR (dávka L), byly hladiny proteinu AChE v kůře výrazně sníženy ve srovnání se skupinou vyvolanou skopolaminem (obr. 6C, F(8,18) = 50. Obr. 6C.590, P < 0,001; F(8,18) = 16,950, P < 0,05; F(8,18) = 30,993, P < 0,01; F(8,18) = 23,821, P < 0.01; F(8,18) = 52,196, P < 0,001; F(8,18) = 29,284, P < 0,01), zatímco hladiny ChAT nebyly změněny (obr. 6E). A podobné účinky na hladiny AChE a ChAT byly pozorovány při podávání HAL a HAR (dávky L, M a H) v hipokampu (obr. 6D, F(8,18) = 41,195, P < 0.001; F(8,18) = 28,111, P < 0,01; F(8,18) = 47,611, P < 0,001; F(8,18) = 30,539, P < 0,01; F(8,18) = 33,225, P < 0,01; F(8,18) = 40,032, P < 0,001; obrázek 6F). Kromě toho byl inhibiční účinek HAR na expresi proteinu AChE závislý na dávce, zatímco HAL měl výrazný inhibiční účinek při nízké dávce 2 mg/kg (obr. 6). Léčba HAL a HAR tedy zabránila skopolaminem indukovanému zvýšení aktivity AChE a exprese proteinu a snížení aktivity ChAT, což prokázalo, že HAL a HAR mohou zlepšit funkci cholinergního systému a zmírnit behaviorální dysfunkci. Zejména u HAL se projevilo významné zlepšení při nízkém dávkování.
Vliv HAL a HAR na markery oxidačního stresu
Měřily se účinky HAL a HAR na aktivity antioxidačních enzymů SOD, GSH-px a hladinu MDA v mozkové kůře. Jak ukázal obrázek 5, u skupiny navozené skopolaminem došlo ve srovnání s kontrolní skupinou k viditelnému útlumu aktivit SOD a GSH-px (obrázek 5A, F(8,81) = 12,254, P < 0,01; obrázek 5B, F(8,81) = 4,577, P < 0,05). Zatímco podávání HAL (dávka H), HAR (dávky M a H) a donepezilu významně zvýšilo aktivitu SOD (obr. 5A, F(8,81) = 13,287, P < 0,01; F(8,81) = 13,424, P < 0,01; F(8,81) = 34,510, P < 0,01; F(8,81) = 34,510, P < 0,01).001; F(8,81) = 8,670, P < 0,01) a léčba HAL, HAR (obě dávky H) a donepezilem jasně zvýšila aktivitu GSH-px (obr. 5B, F(8,81) = 6,010, P < 0,05; F(8,81) = 6,008, P < 0,05; F(8,81) = 6,860, P < 0,05). Dále bylo při léčbě HAL, HAR (dávka M i H) a donepezilem (5 mg/kg) pozorováno významné zmírnění zvýšení hladiny MDA ve srovnání s myší vyvolanou skopolaminem (obr. 5C, F(8,81) = 9.302, P < 0,01; F(8,81) = 6,565, P < 0,05; F(8,81) = 12,690, P < 0,01; F(8,81) = 33,627, P < 0,001; F(8,81) = 5,874, P < 0,05). Tyto výsledky naznačují, že HAL a HAR mohou bránit oxidačnímu poškození vyvolanému skopolaminem prostřednictvím ochrany kůry před oxidačním stresem. Je pozoruhodné, že oxidační poškození bylo možné zlepšit pouze při vysokých dávkách jak u HAL, tak u HAR a zlepšení u HAL a HAR bylo obecně srovnatelné.
Vliv HAL a HAR na hladiny zánětu
Byly analyzovány účinky HAL a HAR na aktivity zánětlivých faktorů MPO, TNF-α, IL-1β, IL-6, IL-10 a hladiny NO v kůře nebo hipokampu. Aktivity MPO (kůra a hipokampus) a TNF-α (kůra) byly u myší indukovaných skopolaminem zvýšené v porovnání s kontrolními myšmi (obr. 4C, F(8,81) = 34.).740, P < 0,001; obrázek 4F, F(8,81) = 6,729, P < 0,05; obrázek 5D, F(8,81) = 6,316, P < 0,05), ale hladiny IL-1β, IL-6 a IL-10 se nezměnily (obrázky 5F-H). Podávání HAL, HAR (dávky M i H) a donepezilu však vedlo k významnému snížení aktivity MPO v kůře a hipokampu v závislosti na dávce (obr. 4C, F(8,81) = 10.435, P < 0,01; F(8,81) = 32,309, P < 0,001; F(8,81) = 16,397, P < 0,01; F(8,81) = 31,291, P < 0.001; F(8,81) = 7,129, P < 0,05; obrázek 4F, F(8,81) = 7,212, P < 0,05; F(8,81) = 10,533, P < 0.01; F(8,81) = 9,150, P < 0,01; F(8,81) = 18,903, P < 0,001; F(8,81) = 11,641, P < 0,01), a léčba HAL (dávky L, M a H), HAR (dávky M a H) a donepezilem rovněž oslabuje aktivitu TNF-α v kůře (obr. 5D, F(8,81) = 10.).377, P < 0,01; F(8,81) = 28,257, P < 0,001; F(8,81) = 31,088, P < 0,001; F(8,81) = 15,966, P < 0,001; F(8,81) = 23,530, P < 0,001; F(8,81) = 5,909, P < 0,05). Kromě skupiny s dávkováním HAR-L byl navíc u ostatních myší léčených HAL, HAR a donepezilem zjištěn znatelný útlum zvýšení hladiny NO ve srovnání s myší s amnézií vyvolanou skopolaminem (obr. 5E, F(8,81) = 21.).319, P < 0,001; F(8,81) = 12,996, P < 0,01; F(8,81) = 11,911, P < 0,01; F(8,81) = 5,227, P < 0,05; F(8,81) = 8,799, P < 0.01; F(8,81) = 17,349, P < 0,001) a inhibiční účinky HAL na TNF-α a NO byly výraznější než účinky HAR.
Jak ukazuje obrázek 7, ve srovnání s kontrolními myšmi byly hladiny exprese proteinů kortikální a hipokampální MPO u myší indukovaných skopolaminem zvýšené (obrázek 7C, F(8,18) = 29,975, P < 0,01; obrázek 7D, F(8,18) = 32,014, P < 0,01). Po podání donepezilu, HAL a HAR (dávky L, M a H) exprese proteinů MPO v kůře a hipokampu zjevně poklesla ve srovnání se skupinou vyvolanou skopolaminem. Kromě toho výsledky imunofluorescenčního stanovení také odhalily, že léčba HAL a HAR mohla nápadně snížit expresi MPO v kůře v závislosti na dávce a inhibice HAL a HAR byly srovnatelné (obr. 8). Všechna tato zjištění naznačují, že HAL a HAR by mohly zmírnit zánětlivé poškození vyvolané skopolaminem prostřednictvím inhibice zánětu v kůře a hipokampu.
Modulační účinky HAL a HAR na neurotransmitery myší léčených skopolaminem
Různé plazmatické a kortikální neurotransmitery byly hodnoceny pomocí UPLC-ESI-MS/MS a změny jsou uvedeny na obrázcích 9, resp. 10. Na obrázcích 9 a 10 jsou znázorněny změny v neurotransmiterech. Na základě dříve zavedené metody stanovení neurotransmiterů byly obsahy ACh, Ch, L-Trp, 5-HT, L-Glu, L-Phe, L-Tyr v plazmě a ACh, Ch, L-Trp, γ-GABA, L-Glu, L-Phe, L-Tyr v kůře nad dolní mezí stanovitelnosti a všechny pak byly úspěšně kvantifikovány.
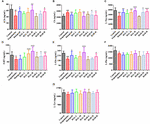
Obrázek 9. Účinky HAL (2, 5 a 10 mg/kg, konkrétně nízké: L, střední: M a vysoké: H dávky) a HAR (10, 20 a 30 mg/kg, konkrétně L, M a H dávky) na obsah neurotransmiterů v plazmě myší s kognitivními poruchami vyvolanými skopolaminem. N = 10/skupina. ACh (A), Ch (B), L-Trp (C), 5-HT (D), L-Glu (E), L-Phe (F), L-Tyr (G). #P < 0,05; ##P < 0,01; ###P < 0,001, oproti kontrolní skupině. ∗P < 0,05; ∗∗P < 0,01; ∗∗∗P < 0,001, oproti skupině navozené skopolaminem.
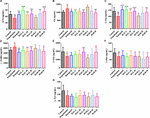
OBRÁZEK 10. Ukazuje, že skupina navozená skopolaminem je velice dobrá. Účinky HAL (2, 5 a 10 mg/kg, konkrétně nízké: L, střední: M a vysoké: H dávky) a HAR (10, 20 a 30 mg/kg, konkrétně L, M a H dávky) na obsah neurotransmiterů v mozkové kůře myší s kognitivními poruchami vyvolanými skopolaminem. N = 10 skupin. ACh (A), Ch (B), L-Trp (C), γ-GABA (D), L-Glu (E), L-Phe (F), L-Tyr (G). #P < 0,05; ##P < 0,01, oproti kontrolní skupině. ∗P < 0,05; ∗∗P < 0,01; ∗∗∗P < 0,001, vs. skupina vyvolaná skopolaminem.
Po indukci skopolaminem v dávce 1 mg/kg došlo k poklesu koncentrací ACh, L-Trp, 5-HT, L-Glu a L-Phe v plazmě (obr. 9A, F(8,81) = 11.).453, P < 0,01; obrázek 9C, F(8,81) = 23,137, P < 0,001; obrázek 9D, F(8,81) = 7,359, P < 0,05; obrázek 9E, F(8,81) = 55.531, P < 0,001; obrázek 9F, F(8,81) = 6,330, P < 0,05), zatímco bez vlivu na L-Tyr (obrázek 9G, F(8,81) = 2,091, P > 0,05). Při ošetření HAL a HAR ve vysokých dávkách však došlo ke zvýšení obsahu ACh a L-Glu (obr. 9A,E) a obsah L-Trp byl zjevně zvýšen při podávání HAL (dávky L, M a H) a HAR (dávky M a H) v závislosti na dávce (obr. 9C). Myši léčené HAL a HAR (dávky M a H) navíc vykazovaly významné zvýšení hladiny 5-HT ve srovnání s myšmi, kterým byl podáván skopolamin (obr. 9D). Podávání HAL a HAR však nemodulovalo obsah L-Phe a L-Tyr (obr. 9F,G). Kromě toho HAL (dávka H) i HAR (dávky M a H) vyvolaly účinek na snížení obsahu Ch v plazmě (obrázek 9B).
Obrázek 10 dále ukazuje změny neurotransmiterů v mozkové kůře. Po indukci skopolaminem se zjevně snížil obsah ACh, L-Trp, L-Glu, L-Phe a L-Tyr (Obrázek 10A, F(8,81) = 11,296, P < 0,01; Obrázek 10C, F(8,81) = 10,979, P < 0,01; Obrázek 10E, F(8,81) = 7,216, P < 0,05; Obrázek 10F, F(8,81) = 10.340, P < 0,01; obrázek 10G, F(8,81) = 6,834, P < 0,05), zatímco obsah Ch a γ-GABA byl v kůře významně zvýšen ve srovnání s kontrolní skupinou (obrázek 10B, F(8,81) = 7,377, P < 0,05; obrázek 10D, F(8,81) = 5,580, P < 0,05). Nicméně obsah ACh byl pozorovatelně zvýšen ve skupinách HAL (dávky L, M a H) a HAR (dávky M a H) a obsah L-Trp byl nápadně zvýšen ve skupinách HAL (dávky L, M a H) a HAR (dávka H) a množství L-Glu bylo rovněž zvýšeno u vysokých dávek skupin HAL a HAR (obr. 10A,C,E). Kromě toho léčba HAL (dávky L, M a H) a HAR (dávka H) vedla ke zjevnému snížení hladin Ch a γ-GABA ve srovnání se skupinou vyvolanou skopolaminem (obr. 10B,D). Různé dávky HAL a HAR však vykazovaly zanedbatelný vliv na pokles L-Phe a L-Tyr vyvolaný skopolaminem (obr. 10F,G). Obecně byly účinky HAL na důležité plazmatické a kortikální neurotransmitery výraznější než účinky HAR (obr. 9, 10).
Diskuse
Alzheimerova choroba je smrtelně nebezpečné progresivní neurodegenerativní onemocnění s rostoucím počtem stárnoucí populace a prodlužující se délkou života, a proto je naléhavě nutné najít nové léky, které by mohly AD účinně léčit (Kouémou et al., 2017). Je obecně známo, že cholinergní hypofunkce, oxidační stres a neurozánět jsou jednou z hlavních charakteristik AD (Abd-El-Fattah et al., 2014; Demirci et al., 2017). Skopolamin je neselektivní antagonista muskarinového cholinergního receptoru (Jang et al., 2013; Kaur et al., 2017). Četné studie ukázaly, že skopolamin může zhoršovat schopnost učení a paměti u lidí i na zvířecích modelech, a model AD vyvolaný skopolaminem se široce používá ke studiu kognitivního deficitu a ke screeningu léčiv proti AD (Goverdhan et al., 2012). Skopolamin vede ke kognitivnímu deficitu spojenému se snížením cholinergní neurotransmise, zvýšeným oxidačním stresem i zvýšeným zánětem v mozku (Lin a Beal, 2006). Léčiva proto mohou účinně zvýšit cholinergní funkci a/nebo zmírnit oxidační stres a zánět a mohou zvrátit poruchy učení a paměti u myší s AD vyvolanou skopolaminem a v konečném důsledku budou cenná pro léčbu AD.
Předtím He et al. (2015a) prokázali, že HAR může inhibovat aktivitu AChE a významně zlepšit prostorové učení a paměť kognitivně postižených myší, včetně transgenních myší s APP/PS1 a myší s AD vyvolanou skopolaminem. V této studii byly porovnány paměť zlepšující účinky HAL (2, 5 a 10 mg/kg) a HAR (10, 20 a 30 mg/kg) na myši indukované skopolaminem pomocí hodnocení chování MWM. Bylo ověřeno, že skopolamin (1 mg/kg) poškozuje schopnost prostorového učení a paměti myší v úloze MWM, což bylo v souladu s našimi předchozími studiemi (He et al., 2015a; Liu et al., 2017). Po léčbě přípravky HAL (dávky L, M a H) a HAR (dávky M a H) se snížila latence útěku a délka cesty a účinně se zvýšila frekvence procházení (obr. 2, 3), což ukazuje, že podávání přípravků HAL a HAR znatelně zlepšilo učení a prostorovou paměť myší vyvolané skopolaminem a zlepšení u HAL bylo výraznější než u HAR.
Pro další objasnění základního mechanismu zlepšení učení a paměti u myší indukovaných skopolaminem byly hodnoceny změny cholinergního systému, oxidačního stresu a zánětu. Jak bylo uvedeno v předchozí literatuře (Abd-El-Fattah et al., 2014; Lin et al., 2016; Demirci et al., 2017), v současné studii vedla i.p. injekce skopolaminu k útlumu cholinergní funkce, zvýšenému oxidačnímu stresu a zánětu. Donepezil je jako inhibitor AChE reprezentativní symptomatickou léčbou mírné a středně těžké formy AD (Bohnen et al., 2005) a nedávné výzkumy odhalily, že může také potlačovat zánět a oxidační stres (Saxena et al., 2008; Yoshiyama et al., 2010). V této studii mohl donepezil (pozitivní kontrola) účinně zmírnit tyto změny u myší vyvolané skopolaminem a zlepšit jejich učení a paměť. Podobně jako jejich aktivity in vitro (Zhao et al., 2013; Liu et al., 2014), HAL a HAR v různých dávkách nejen zvrátily skopolaminem indukované zvýšení aktivity AChE, ale také vedly ke zvýšení neurotransmiteru ACh a snížení Ch v mozkové kůře, hipokampu nebo plazmě (obr. 4, 9, 10). HAL a HAR navíc zjevně inhibovaly hladiny proteinu kortikální AChE (obr. 6C,D). Jak uvádí Oh et al. (2009), ChAT-pozitivní neurony byly významně redukovány léčbou skopolaminem. Současné výsledky ukázaly, že HAL a HAR mohou zvrátit snížení aktivity ChAT vyvolané skopolaminem (obr. 4B,E), přičemž nemají žádný vliv na expresi proteinu ChAT (obr. 6E,F), a to je třeba dále prokázat. Tím se tedy potvrdilo, že anti-amnestické účinky HAL a HAR na poruchy poznávacích funkcí vyvolané skopolaminem pravděpodobně souvisejí s modifikací cholinergního systému (zejména AChE) a HAL prokázal zjevné cholinergní zlepšení při nízkém dávkování (2 mg/kg).
Mnoho studií navíc ukázalo, že zvýšený oxidační stres je široce považován za významný přispívající rizikový faktor při rozvoji AD (Ghumatkar et al., 2015; Lu et al., 2018). HAL a HAR mohou sloužit jako endogenní antioxidanty a bylo zjištěno, že mají antioxidační kapacitu tím, že vychytávají volné radikály (Tse-Susanna et al., 1991; Berrougui et al., 2006a,b). V této studii byla u myší indukovaných skopolaminem pozorována nadprodukce ROS (produkt peroxidace lipidů, MDA) a snížení antioxidačních enzymů (SOD a GSH-px) (obr. 5). Podávání HAL a HAR však pozorovatelně aktivovalo antioxidační enzymy SOD a GSH-px a potlačilo tvorbu MDA v kůře myší s demencí vyvolanou skopolaminem (obr. 5) a poté zlepšilo schopnost antioxidační obrany, což svědčí o neuroprotektivních účincích HAL a HAR. Pozoruhodné je, že antioxidační účinky HAL a HAR byly obecně srovnatelné.
Navíc stále více důkazů naznačuje, že zhoršení paměti je spojeno se zvýšeným neurozápalem (Abd-El-Fattah et al., 2014). Podle studie Bensalema et al. (2014) vykazovaly HAL a HAR významnou inhibici MPO s IC50 0,08 a 0,26 μM. Protizánětlivé aktivity HAR bylo dosaženo prostřednictvím inhibice TNF-α a NO u myších buněk RAW264 a lidských THP-1 stimulovaných lipopolysacharidem (Patel et al., 2012). Pozoruhodné je, že v mozku myší s amnézií vyvolanou skopolaminem byly zjištěny vysoké hladiny prozánětlivých cytokinů (TNF-α a MPO) a mediátoru zánětu (NO) (obr. 4, 5). Současná studie potvrdila, že léčba HAL a HAR může současně inhibovat tvorbu NO a upregulaci TNF-α a MPO, přičemž inhibiční účinky HAL na TNF-α a NO byly zřejmější než účinky HAR (obr. 4, 5). Kromě toho výsledky western blottingu a imunofluorescenční analýzy dále dokazují, že hladiny exprese proteinu MPO byly výrazně potlačeny HAL a HAR (obrázky 7, 8), což naznačuje, že schopnost HAL a HAR působit proti paměťovým deficitům může být zprostředkována potlačením zánětlivých markerů TNF-α, MPO a NO.
Jak je známo, AD je spojena s nedostatečnou hladinou různých neurotransmiterů (Kumar a Singh, 2015; Svob Strac et al., 2015). Předchozí studie uváděly, že myši, kterým byl podáván samotný skopolamin, vykazovaly značně sníženou reaktivitu cholinergního systému, což naznačovala snížená hladina ACh a zvýšená aktivita AChE spolu se sníženou aktivitou ChAT (Lee et al., 2014). Transgenní zvířecí model AD prokázal pokles hladiny Glu (Nilsen et al., 2012) a sérová a plazmatická metabolomika rovněž naznačila pokles Trp, 5-HT, Glu, Fe, Tyr u myší nebo pacientů s modelem AD (Vermeiren et al., 2014; González-Domínguez et al., 2015; Weng et al., 2015; Corso et al., 2017; Li et al., 2018). V souladu s tím naše výsledky prokázaly, že demence vyvolaná skopolaminem byla doprovázena poklesem ACh, L-Trp, 5-HT, L-Glu, L-Phe, L-Tyr a zvýšením Ch a γ-GABA, což je v souladu s předchozími studiemi (Scali et al., 1999; Nilsen et al., 2012; Lee et al., 2014; Vermeiren et al., 2014; González-Domínguez et al., 2015; Kumar a Singh, 2015; Weng et al., 2015; Zhou et al., 2016; Corso et al., 2017; Liu et al., 2017). Neurotransmitery hrají důležitou roli v mozkovém okruhu zapojeném do mnoha aspektů učení a paměti, zejména cholinergní, serotonergní, glutametargické a GABAergní neurotransmitery (Vermeiren et al., 2014; Kumar a Singh, 2015; Zhou et al., 2016). Je pozoruhodné, že s výjimkou L-Phe a L-Tyr bylo možné ostatní změny účinně zvrátit podáním HAL i HAR v plazmě a kůře, přičemž modulace HAL byly výraznější než modulace HAR (obr. 9, 10). Regulace těchto neurotransmiterů pomocí HAL a HAR tedy může být prospěšná pro zlepšení schopnosti učení a paměti myší s demencí.
Všeobecně řečeno, HAL (2, 5 a 10 mg/kg) a HAR (20 a 30 mg/kg) vykazovaly zvýšení cholinergní neurotransmise prostřednictvím potlačení AChE a indukce ChAT, zvýšení antioxidační ochrany zvýšením aktivity SOD a GSH-px a snížením produkce MDA, zvýšení protizánětlivých účinků snížením TNF-α, MPO a NO a modulací hladin kritických neurotransmiterů (ACh, Ch, L-Trp, 5-HT, γ-GABA a L-Glu). HAL a HAR tak mohou působit na více cílů jako terapeutika pro AD. Kromě toho byly regulace HAL na cholinergní funkci, zánět a neurotransmitery výraznější než regulace HAR a HAL vykazoval podobnou antioxidační kapacitu jako HAR. Zejména účinná dávka HAL (2 mg/kg) byla mnohem nižší než dávka HAR (20 mg/kg), což může být způsobeno zejména rozdíly v biologické dostupnosti a metabolické stabilitě obou léčiv (Li et al., 2017b). Souhrnně všechny tyto výsledky ukázaly, že ve srovnání s HAR může být HAL vhodnější kandidátskou sloučeninou s lepšími anti-amnestickými a farmakokinetickými vlastnostmi pro terapii AD.
Závěr
Shrnem lze říci, že HAL a HAR by mohly účinně zmírnit poruchy paměti na myším modelu vyvolaném skopolaminem prostřednictvím zlepšení funkce cholinergního systému, potlačení oxidačního stresu a poškození zánětu a modulace životně důležitých neurotransmiterů u myší s demencí. Zejména u HAL byla regulace cholinergní funkce, zánětu a neurotransmiterů významnější než u HAR a vykazovala srovnatelný antioxidační účinek jako HAR. Pozoruhodné je, že účinná dávka HAL (2 mg/kg) byla mnohem nižší než dávka HAR (20 mg/kg). Celkové výsledky naznačují, že HAL může být lepším kandidátským léčivem pro léčbu poruch učení a paměti, jako je AD. Je třeba provést další hodnocení anti-amnestických účinků a molekulárních mechanismů HAL, což poskytne podstatné počáteční důkazy pro prevenci a léčbu AD.
Příspěvky autorů
S-PL a C-HW: podíleli se na návrhu výzkumu, provedli analýzu dat, napsali rukopis nebo se na něm podíleli. S-PL, Y-WW, S-LQ, Y-PZ, GD, W-ZD, CM, Q-YL, H-DG, WL a X-MC: prováděli experimenty.
Financování
Tato práce byla podpořena Čínskou národní nadací pro přírodní vědy a Ujgurskou autonomní oblastí Sin-ťiang (grant č. 1). U1130303), National Natural Science Foundation of China (Grant No. 81173119), Key Project of Ministry of Science and Technology of China (Grant No. 2012ZX09103201-051) a Projekty technologické spolupráce vědy v Šanghaji v Číně (grant č. 14495800200) udělené C-HW za finanční podporu této studie.
Prohlášení o střetu zájmů
Autoři prohlašují, že výzkum byl prováděn bez jakýchkoli komerčních nebo finančních vztahů, které by mohly být chápány jako potenciální střet zájmů.
Abd-El-Fattah, M. A., Abdelakader, N. F., a Zaki, H. F. (2014). Pyrrolidin dithiokarbamát chrání před kognitivními poruchami vyvolanými skopolaminem u potkanů. Eur. J. Pharmacol. 723, 330-338. doi: 10.1016/j.ejphar.2013.11.008
PubMed Abstract | CrossRef Full Text | Google Scholar
Aucoin, J. S., Jiang, P., Aznavour, N., Tong, X. K., Buttini, M., Descarries, L., et al. (2005). Selektivní cholinergní denervace nezávislá na oxidačním stresu v myším modelu Alzheimerovy choroby. Neuroscience 132, 73-86. doi: 10.1016/j.neuroscience.2004.11.047
PubMed Abstract | CrossRef Full Text | Google Scholar
Bassani, T. B., Turnes, J. M., Moura, E. L., Bonato, J. M., Cóppola-Segovia, V., Zanata, S. M., et al. (2017). Účinky kurkuminu na krátkodobou prostorovou a rozpoznávací paměť, neurogenezi dospělých a neurozánět u streptozotocinem indukovaného modelu demence Alzheimerova typu u potkanů. Behavior. Brain Res. 335, 41-54. doi: 10.1016/j.bbr.2017.08.014
PubMed Abstract | CrossRef Full Text | Google Scholar
Bensalem, S., Soubhye, J., Aldib, I., Bournine, L., Nguyen, A. T., Vanhaeverbeek, M., et al. (2014). Inhibice aktivity myeloperoxidázy alkaloidy Peganum harmala L. (Zygophyllaceae). J. Ethnopharmacol. 154, 361-369. doi: 10.1016/j.jep.2014.03.070
PubMed Abstract | CrossRef Full Text | Google Scholar
Berrougui, H., Isabelle, M., Cloutier, M., Hmamouchi, M., and Khalil, A. (2006a). Ochranné účinky extraktu z Peganum harmala L., harminu a harmalinu proti oxidaci lidského lipoproteinu o nízké hustotě. J. Pharm. Pharmacol. 58, 967-974. doi: 10.1211/jpp.58.7.0012
PubMed Abstract | CrossRef Full Text | Google Scholar
Berrougui, H., Martín-Cordero, C., Khalil, A., Hmamouchi, M., Ettaib, A., Marhuenda, E., et al. (2006b). Vasorelaxační účinky harminu a harmalinu extrahovaných ze semen Peganum harmala L. v izolované aortě potkana. Pharmacol. Res. 54, 150-157. doi: 10.1016/j.phrs.2006.04.001
PubMed Abstract | CrossRef Full Text | Google Scholar
Bohnen, N. I., Kaufer, D. I., Hendrickson, R., Ivanco, L. S., Lopresti, B. J., Koeppe, R. A., et al. (2005). Stupeň inhibice aktivity kortikální acetylcholinesterázy a kognitivní účinky léčby donepezilem u Alzheimerovy choroby. J. Neurol. Neurosurg. Psychiatry 76, 315-319. doi: 10.1136/jnnp.2004.038729
PubMed Abstract | CrossRef Full Text | Google Scholar
Bromley-Brits, K., Deng, Y., and Song, W. H. (2011). Morrisův test vodního bludiště pro zjišťování deficitů učení a paměti u myší na modelu Alzheimerovy choroby. J. Vis. Exp. 53, 1-5. doi: 10.3791/2920
PubMed Abstract | CrossRef Full Text | Google Scholar
Corso, G., Cristofano, A., Sapere, N., La Marca, G., Angiolillo, A., Vitale, M., et al. (2017). Sérové profily aminokyselin u normálních osob a u pacientů s Alzheimerovou demencí nebo s rizikem jejího vzniku. Dement. Geriatr. Cogn. Dis. Extra 7, 143-159. doi: 10.1159/000466688
PubMed Abstract | CrossRef Full Text | Google Scholar
Coyle, J. T., Price, D. L., and Delong, M. R. (1983). Alzheimerova choroba: porucha kortikální cholinergní inervace. Science 219, 1184-1190. doi: 10.1126/science.6338589
CrossRef Full Text | Google Scholar
Demirci, K., Nazıroğlu, M., Övey,İ. S., and Balaban, H. (2017). Selen tlumí apoptózu, zánět a oxidační stres v krvi a mozku starých potkanů s demencí vyvolanou skopolaminem. Metab. Brain Dis. 32, 321-329. doi: 10.1007/s11011-016-9903-1
PubMed Abstract | CrossRef Full Text | Google Scholar
Fan, L. Y., and Chiu, M. J. (2014). Kombinovaná terapie a současné koncepty i budoucí strategie léčby Alzheimerovy choroby. Neuropsychiatr. Dis. Treat. 10, 439-451. doi: 10.2147/NDT.S45143
PubMed Abstract | CrossRef Full Text | Google Scholar
Frison, G., Favretto, D., Zancanaro, F., Fazzin, G., and Ferrara, S. D. (2008). Případ intoxikace beta-karbolinovými alkaloidy po požití extraktu ze semen Peganum harmala. Forensic Sci. Int. 179, 37-43. doi: 10.1016/j.forsciint.2008.05.003
PubMed Abstract | CrossRef Full Text | Google Scholar
Ghumatkar, P. J., Patil, S. P., Jain, P. D., Tambe, R. M., and Sathaye, S. (2015). Nootropní, neuroprotektivní a neurotrofní účinky floretinu u skopolaminem indukované amnézie u myší. Pharmacol. Biochem. Behavior. 135, 182-191. doi: 10.1016/j.pbb.2015.06.005
PubMed Abstract | CrossRef Full Text | Google Scholar
González-Domínguez, R., García-Barrera, T., and Gómez-Ariza, J. L. (2015). Profilování metabolitů pro identifikaci změněných metabolických drah u Alzheimerovy choroby. J. Pharm. Biomed. Anal. 107, 75-81. doi: 10.1016/j.jpba.2014.10.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Goverdhan, P., Sravanthi, A., and Mamatha, T. (2012). Neuroprotektivní účinky meloxikamu a selegilinu u kognitivních poruch vyvolaných skopolaminem a oxidačním stresem. Int. J. Alzheimers Dis. 2012, 1-8. doi: 10.1155/2012/974013
PubMed Abstract | CrossRef Full Text | Google Scholar
Gray, E., Thomas, T. L., Betmouni, S., Betmouni, S., Scolding, N., and Love, S. (2008). Zvýšená aktivita a mikrogliální exprese myeloperoxidázy v demyelinizované mozkové kůře u roztroušené sklerózy. Brain Pathol. 18, 86-95. doi: 10.1111/j.1750-3639.2007.00110.x
PubMed Abstract | CrossRef Full Text | Google Scholar
He, D. D., Wu, H., Wei, Y., Liu, W., Huang, F., Shi, H. L., et al. (2015a). Účinky harminu, inhibitoru acetylcholinesterázy, na prostorové učení a paměť transgenních myší APP/PS1 a myší s poruchou paměti vyvolanou skopolaminem. Eur. J. Pharmacol. 768, 96-107. doi: 10.1016/j.ejphar.2015.10.037
PubMed Abstract | CrossRef Full Text | Google Scholar
He, D. D., Zhang, L., Liu, L., Wu, X. J., Cheng, X. M., and Wang, C. H. (2015b). Celkové alkaloidy ze semen Peganum harmala zlepšující schopnost učení a paměť myší. Chin. Tradit. Pat. Med. 37, 478-482.
Heneka, M. T., Carson, M. J., El Khoury, J., Landreth, G. E., Brosseron, F., Feinstein, D. L., et al. (2015). Neuroinflammation in Alzheimer’s disease (Neurozánět u Alzheimerovy choroby). Lancet Neurol. 14, 388-405. doi: 10.1016/S1474-4422(15)70016-5
CrossRef Full Text | Google Scholar
Herraiz, T., Gonzalez, D., Ancin-Azpilicueta, C., Arán, V. J., and Guillén, H. (2010). Beta-karbolinové alkaloidy v Peganum harmala a inhibice lidské monoaminooxidázy (MAO). Food Chem. Toxicol. 48, 839-845. doi: 10.1016/j.fct.2009.12.019
PubMed Abstract | CrossRef Full Text | Google Scholar
Jang, Y. J., Kim, J., Shim, J., Kim, C. Y., Jang, J. H., Lee, K. W., et al. (2013). Káva bez kofeinu zabraňuje poruchám paměti vyvolaným skopolaminem u potkanů. Behavior. Brain Res. 245, 113-119. doi: 10.1016/j.bbr.2013.02.003
PubMed Abstract | CrossRef Full Text | Google Scholar
Jeon, S. J., Kim, B., Park, H. J., Zhang, J. B., Kwon, Y., Kim, D. H., et al. (2017). Zlepšující účinek 1-palmitoyl-2-linoleoyl-3-acetylglycerolu na poruchy paměti vyvolané skopolaminem prostřednictvím inhibice acetylcholinesterázy a aktivace LTP. Behavior. Brain Res. 324, 58-65. doi: 10.1016/j.bbr.2017.01.040
PubMed Abstract | CrossRef Full Text | Google Scholar
Jiang, B. (2016). Study on the Transportation of Alkaloids from Peganum harmala through the Blood Brain Barrier and the Mechanism of Their Effect on the Neurotransmitters in Rats Brain (Studium transportu alkaloidů z Peganum harmala přes hematoencefalickou bariéru a mechanismu jejich účinku na neurotransmitery v mozku potkanů). Doktorská disertační práce, Shanghai University of Traditional Chinese Medicine, Shanghai.
Jiménez, J., Riveron-Negrete, L., Abdullaev, F., Espinosa-Aguirrec, J., and Rodríguez-Arnaiza, R. (2008). Cytotoxicita β-karbolinových alkaloidů harminu a harmalinu v testech na lidských buňkách in vitro. Exp. Toxicol. Pathol. 60, 381-389. doi: 10.1016/j.etp.2007.12.003
PubMed Abstract | CrossRef Full Text | Google Scholar
Kaur, R., Singh, V., and Shri, R. (2017). Antiamnestické účinky druhů Ganoderma: možný cholinergní a antioxidační mechanismus. Biomed. Pharmacother. 92, 1055-1061. doi: 10.1016/j.biopha.2017.06.029
PubMed Abstract | CrossRef Full Text | Google Scholar
Kouémou, N. E., Taiwe, G. S., Moto, F. C. O., Pale, S., Gwladys, N. T., Njapdounke, J. S. K., et al. (2017). Nootropní a neuroprotektivní účinky dichrocephala integrifolia na skopolaminovém myším modelu Alzheimerovy choroby. Front. Pharmacol. 8:847. doi: 10.3389/fphar.2017.00847
CrossRef Full Text | Google Scholar
Kumar, A., and Singh, A. (2015). Přehled patofyziologie Alzheimerovy choroby a její léčby: aktualizace. Pharmacol. Rep. 67, 195-203. doi: 10.1016/j.pharep.2014.09.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Kwon, S. H., Lee, H. K., Kim, J. A., Hong, S. I., Kim, H. C., Jo, T. H., et al. (2010). Neuroprotektivní účinky kyseliny chlorogenové na amnézii vyvolanou skopolaminem prostřednictvím antiacetylcholinesterázy a antioxidačních aktivit u myší. Eur. J. Pharmacol. 649, 210-217. doi: 10.1016/j.ejphar.2010.09.001
PubMed Abstract | CrossRef Full Text | Google Scholar
Lee, H. E., Jeon, S. J., Ryu, B., Park, S. J., Ko, S. Y., Lee, Y., et al. (2016). Swertisin, C-glukosylflavon, zlepšuje poruchy paměti vyvolané skopolaminem u myší díky své antagonistické vlastnosti adenosinového receptoru A1. Behavior. Brain Res. 306, 137-145. doi: 10.1016/j.bbr.2016.03.030
PubMed Abstract | CrossRef Full Text | Google Scholar
Lee, S., Kim, J., Seo, S. G., Choi, B. R., Han, J. S., Lee, K. W., et al. (2014). Sulforafan zmírňuje poruchy paměti vyvolané skopolaminem u myší. Pharmacol. Res. 85, 23-32. doi: 10.1016/j.phrs.2014.05.003
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, J., Liu, Y., Li, W., Wang, Z., Guo, P., Li, L., et al. (2018). Metabolické profilování účinků ginsenosidu Re na myším modelu Alzheimerovy choroby. Behavior. Brain Res. 337, 160-172. doi: 10.1016/j.bbr.2017.09.027
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, S. P., Cheng, X. M., and Wang, C. H. (2017a). Přehled tradičního využití, fytochemie, farmakologie, farmakokinetiky a toxikologie rodu Peganum. J. Ethnopharmacol. 203, 127-162. doi: 10.1016/j.jep.2017.03.049
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, S. P., Zhang, Y. P., Deng, G., Wang, Y. W., Qi, S. L., Cheng, X. M., et al. (2017b). Expoziční charakteristiky analogických β-karbolinových alkaloidů harmalinu a harminu na základě efluxního transportéru multidrug resistance protein 2. Front. Pharmacol. 8:541. doi: 10.3389/fphar.2017.00541
CrossRef Full Text | Google Scholar
Lin, J. J., Huang, L., Yu, J., Xiang, S. Y., Wang, J. L., Zhang, J. R., et al. (2016). Fukoxantin, mořský karotenoid, zvrací kognitivní poruchy vyvolané skopolaminem u myší a inhibuje acetylcholinesterázu in vitro. Mar. Drugs 14:67. doi: 10.3390/md14040067
PubMed Abstract | CrossRef Full Text | Google Scholar
Lin, M. T., and Beal, M. F. (2006). Mitochondriální dysfunkce a oxidační stres u neurodegenerativních onemocnění. Nature 443, 787-795. doi: 10.1038/nature05292
PubMed Abstract | CrossRef Full Text | Google Scholar
Liu, W., Yang, Y. D., Cheng, X. M., Gong, C., Li, S. P., He, D. D. a další (2014). Rychlá a citlivá detekce inhibičních aktivit inhibitorů acetyl- a butyryl-cholinesteras pomocí UPLC-ESIMS/MS. J. Pharm. Biomed. Anal. 94, 215-220. doi: 10.1016/j.jpba.2014.02.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Liu, W., Zhu, Y. D., Wang, Y. L., Qi, S. L., Wang, Y. W., Ma, C., et al. (2017). Antiamnestický účinek extraktu a alkaloidní frakce z nadzemních částí Peganum harmala na deficity paměti vyvolané skopolaminem u myší. J. Ethnopharmacol. 204, 95-106. doi: 10.1016/j.jep.2017.04.019
PubMed Abstract | CrossRef Full Text | Google Scholar
Louis, E. D., Jiang, W., Gerbin, M., Mullaney, M. M., and Zheng, W. (2010). Vztah mezi koncentrací harmanu a harminu v krvi u familiárního esenciálního třesu, sporadického esenciálního třesu a kontrol. Neurotoxicology 31, 674-679. doi: 10.1016/j.neuro.2010.08.003
PubMed Abstract | CrossRef Full Text | Google Scholar
Lu, C., Dong, L. M., Lv, J. W., Wang, Y., Fan, B., Wang, F., et al. (2018). 20(S)-protopanaxadiol (PPD) zmírňuje poruchy paměti vyvolané skopolaminem prostřednictvím regulace cholinergního a antioxidačního systému a exprese Egr-1, c-Fos a c-Jun u myší. Chem. Biol. Interact. 279, 64-72. doi: 10.1016/j.cbi.2017.11.008
PubMed Abstract | CrossRef Full Text | Google Scholar
Mahmoudian, M., Jalipour, H., and Dardashti, P. S. (2002). Toxicita Peganum harmala: přehled a kazuistika. Iran. J. Pharmacol. Ther. 1, 1-4.
Google Scholar
Malik, J., Kaur, J., and Choudhary, S. (2017). Standardizovaný extrakt z Lactuca sativa Linn. a jeho frakcí ruší amnézii vyvolanou skopolaminem u myší: možný cholinergní a antioxidační mechanismus. Nutr. Neurosci. 1-12. doi: 10.1080/1028415X.2017.1291166 .
PubMed Abstract | CrossRef Full Text | Google Scholar
Morris, R. G., Garrud, P., Rawlins, J. A., and O’Keefe, J. (1982). Place navigation impaired in rats with hippocampal lesions (Porucha navigace podle místa u potkanů s lézí hipokampu). Nature 297, 681-683. doi: 10.1038/297681a0
CrossRef Full Text | Google Scholar
Nilsen, L. H., Melø, T. M., Sæther, O., Witter, M. P., and Sonnewald, U. (2012). Změněný neurochemický profil u McGill-R-Thy1-APP modelu Alzheimerovy choroby u potkanů: longitudinální in vivo studie 1H MRS. J. Neurochem. 123, 532-541. doi: 10.1111/jnc.12003
PubMed Abstract | CrossRef Full Text | Google Scholar
Oh, J. H., Choi, B. J., Chang, M. S., and Park, S. K. (2009). Extrakt ze semene Nelumbo nucifera zlepšuje paměť u potkanů s amnézií vyvolanou skopolaminem prostřednictvím indukce exprese cholin acetyltransferázy. Neurosci. Lett. 461, 41-44. doi: 10.1016/j.neulet.2009.05.045
PubMed Abstract | CrossRef Full Text | Google Scholar
Patel, K., Gadewar, M., Tripathi, R., Prasad, S. K., and Patel, D. K. (2012). Přehled léčivého významu, farmakologické aktivity a bioanalytických aspektů beta-karbolinového alkaloidu „Harmine“. Asian Pac. J. Trop. Biomed. 2, 660-664. doi: 10.1016/S2221-1691(12)60116-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Rijpma, A., Meulenbroek, O., and Rikkert, M. O. (2014). Inhibitory cholinesterázy a přídatné doplňky stravy u Alzheimerovy choroby: systematický přehled randomizovaných kontrolovaných studií. Ageing Res. Rev. 16, 105-112. doi: 10.1016/j.arr.2014.06.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Saxena, G., Singh, S. P., Agrawal, R., and Nath, C. (2008). Vliv donepezilu a takrinu na oxidační stres u intracerebrálního modelu demence vyvolaného streptozotocinem u myší. Eur. J. Pharmacol. 581, 283-289. doi: 10.1016/j.ejphar.2007.12.009
PubMed Abstract | CrossRef Full Text | Google Scholar
Scali, C., Prosperi, C., Giovannelli, L., Bianchi, L., Pepeu, G., a Casamenti, F. (1999). β (1-40) Amyloid peptid injekce do nucleus basalis potkanů vyvolává reakci mikroglie a zvyšuje kortikální uvolňování kyseliny γ-aminomáselné in vivo. Brain Res. 831, 319-321. doi: 10.1016/S0006-8993(99)01492-4
CrossRef Full Text | Google Scholar
Shi, X. Y., Liu, W., Zhang, L., Li, S. P., Cheng, X. M., Xi, Y., et al. (2014). Farmakokinetika harmalinu, harminu a jejich metabolitů u potkanů, kterým byly podávány celkové alkaloidní extrakty z Peganum harmala L. Chin. Tradit. Pat. Med. 36, 1169-1175. doi: 10.3969/j.issn.1001-1528.2014.06.013
CrossRef Full Text | Google Scholar
Singh, S., Kaur, H., and Sandhir, R. (2016). Fraktální rozměry: nové paradigma pro hodnocení prostorové paměti a učení pomocí Morrisova vodního bludiště. Behav. Brain Res. 299, 141-146. doi: 10.1016/j.bbr.2015.11.023
PubMed Abstract | CrossRef Full Text | Google Scholar
Svob Strac, D., Muck-Seler, D., and Pivac, N. (2015). Měření neurotransmiterů v mozkomíšním moku pacientů s Alzheimerovou chorobou: přehled. Psychiatr. Danub. 27, 14-24.
Google Scholar
Tse-Susanna, Y. H., Mak, I. T., and Dickens, B. F. (1991). Antioxidační vlastnosti harmanových a β-karbolinových alkaloidů. Biochem. Pharmacol. 42, 459-464. doi: 10.1016/0006-2952(91)90305-O
CrossRef Full Text | Google Scholar
Vermeiren, Y., Van Dam, D., Aerts, T., Engelborghs, S., and De De Deyn, P. P. (2014). Změny monoaminergních neurotransmiterů v postmortálních oblastech mozku depresivních a agresivních pacientů s Alzheimerovou chorobou. Neurobiol. Aging 35, 2691-2700. doi: 10.1016/j.neurobiolaging.2014.05.031
PubMed Abstract | CrossRef Full Text | Google Scholar
Verri, M., Pastoris, O., Dossena, M., Aquilani, R., Guerriero, F., Cuzzoni, G., et al. (2012). Mitochondriální změny, oxidativní stres a neurozánět u Alzheimerovy choroby. Int. J. Immunopathol. Pharmacol. 25, 345-353. doi: 10.1177/039463201202500204
PubMed Abstract | CrossRef Full Text | Google Scholar
Wang, C. H., Zhang, Z. X., Wang, Y. H., and He, X. J. (2015). Cytotoxické indolové alkaloidy proti lidským leukemickým buněčným liniím z toxické rostliny Peganum harmala. Toxins 7, 4507-4518. doi: 10.3390/toxins7114507
PubMed Abstract | CrossRef Full Text | Google Scholar
Weng, R., Shen, S. S., Tian, Y. L., Burton, C., Xu, X. Y., Liu, Y., et al. (2015). Metabolomický přístup odhaluje integrovanou metabolickou síť spojenou s nedostatkem serotoninu. Sci. Rep. 5:11864. doi: 10.1038/srep11864
PubMed Abstract | CrossRef Full Text | Google Scholar
Yoshiyama, Y., Kojima, A., Ishikawa, C., and Arai, K. (2010). Protizánětlivé působení donepezilu zlepšuje tau patologii, synaptickou ztrátu a neurodegeneraci v režimu tauopatie u myší. J. Alzheimers Dis. 22, 295-306. doi: 10.3233/JAD-2010-100681
PubMed Abstract | CrossRef Full Text | Google Scholar
Zhang, L. (2013). Study on the Film Coating Tablets of Total Alkaloids from Peganum harmala [Studie o filmových potahových tabletách celkových alkaloidů z Peganum harmala]. Master Dissertation, Shanghai University of Traditional Chinese Medicine, Shanghai.
Zhao, T., Ding, K. M., Zhang, L., Cheng, X. M., Wang, C. H., and Wang, Z. T. (2013). Inhibiční aktivity β-karbolinových a chinolinových alkaloidních derivátů z rostlin rodu Peganum vůči acetylcholinesteráze a butyrylcholinesteráze. J. Chem. 2013:717232. doi: 10.1155/2013/717232
CrossRef Full Text | Google Scholar
Zhou, X. L., Li, Y. F., Shi, X. Z., and Ma, C. (2016). Přehled terapeutik tlumících hladinu amyloidu β u Alzheimerovy choroby: cílení na neurotransmisi, zánět, oxidační stres a zvýšenou hladinu cholesterolu. Am. J. Transl. Res. 8, 246-269.
PubMed Abstract | Google Scholar
.
Leave a Reply